

Белки являются главным строительным материалом клетки (платическая функция) и контролируют все процессы метаболизма клетки (ферментативная или каталитическая функция). Кроме того, белки выполняют и другие важные функции в клетке: опорно-двигательная (актин, миозин, тубулин), рецепторная, транспортная, регуляторная и т. п.)
По своему строению все белки являются полипептидами и состоят из аминокислот, соединенных пептидными связями (первичная структура белка). Все разнообразие белков определяется аминокис лотным составом, порядком расположения аминокислот и их количеством. Полипептидная цепочка изменяет свое пространственное расположение, закручивается в спираль и удерживается водородны- ми связями (вторичная структура) — такое строение имеют фибрил- лярные белки. Многие белки принимают глобулярную (шаровид- ную) форму за счет образования дополнительных внутримолекулярных связей (сульфидных, других гидрофобных) — так возникает третичная структура белка. А сложные белки (например, гемоглобин) могут включать несколько глобулярных структур (четвертичная структура) и содержать небелковый компонент. Молекула белка способна к упрощению своей организации (от IV→I) и восстановле- нию более высоких уровней при изменении среды (денатурация и ренатурация); если разрушается первичная структура, то ренатурация невозможна (необратимая денатурация).
Нуклеиновые кислоты выполняют генетическую функцию (ДНК) и
участвуют в реализации генетической информации (РНК). Молекулы ДНК и РНК сходны по строению. Это полинуклео- тиды (нуклеотид–мономер), отличающиеся по углеводу (рибоза и дезоксирибоза) и одному из четырех азотистых оснований (Т-ДНК; У-РНК). Кроме того, ДНК состоит из двух полинуклеотидных цепей,
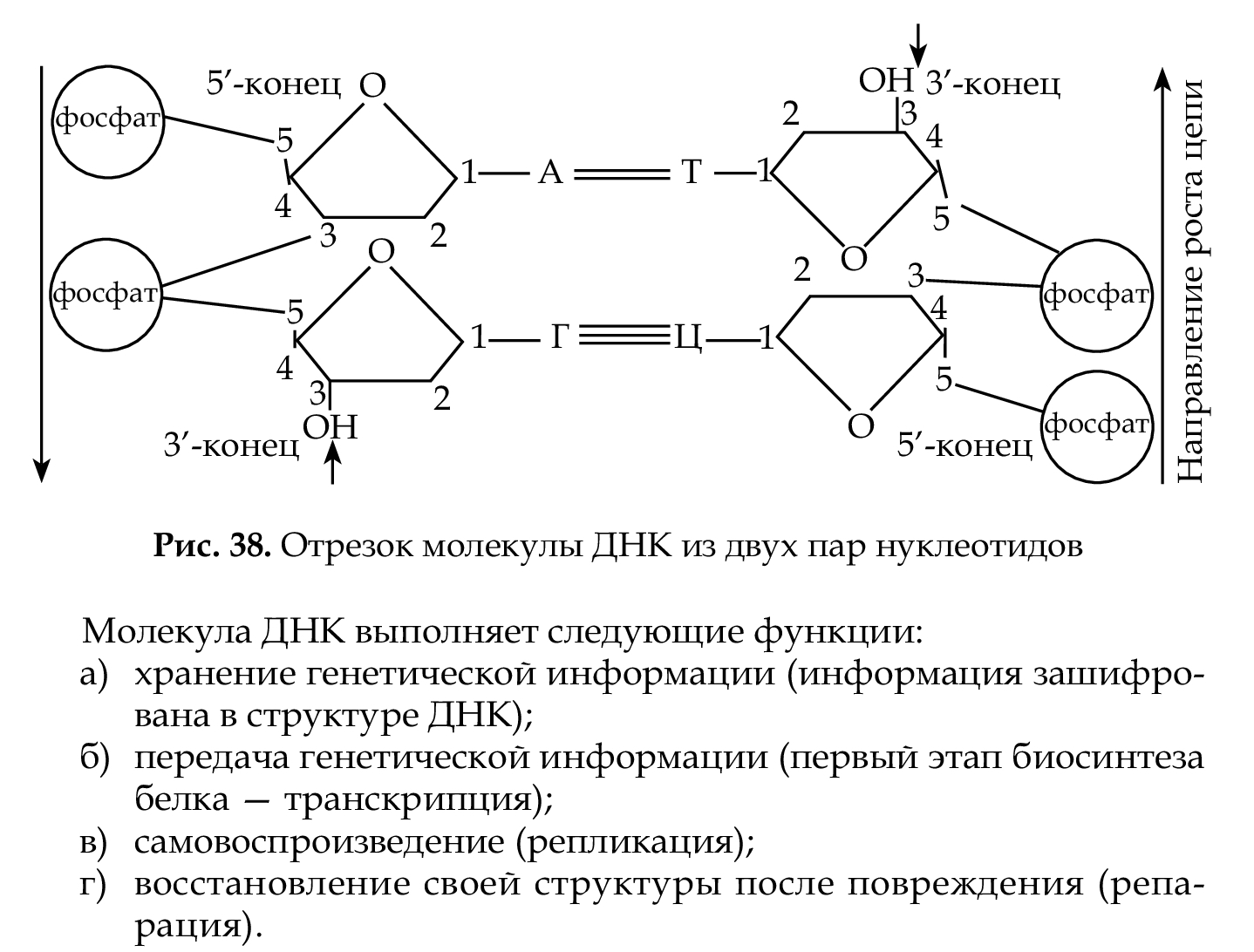
а РНК — из одной. Нуклеотиды одной цепи соединяются связями, возникающими между остатком фосфорной кислоты и углеводом через фосфат, причем углерод в положении 5 последующего нукле- отида соединяется с углеродом в положении 3 предыдущего.
Фермент ДНК-полимераза присоединяет нуклеотиды только в одном направ- лении, а именно: 2-й — к 1-му, 3-й — ко 2-му. Таким образом, удли- нение (рост) цепи идет всегда в одном направлении от 5' → 3', начало цепи обозначается как 5'-конец, а окончание — 3'-конец. В молекуле ДНК (рис. 38) цепи антипараллельны, нуклеотиды противоположных
цепей связаны между собой по правилу комплементарности (А–Т; Г–Ц).
Генетический код — это система записи генетической информа- ции, позволяющая прямой и обратный перевод сведений и ис- пользование их в процессе жизнедеятельности клетки. Свойства генетического кода:
универсальность — это значит, что у всех организмов наслед- ственная информация зашифрована одинаковым способом;
триплетность — единицей генетического кода является три- плет нуклеотидов — кодон, который несет элементарную ин- формацию — об одной аминокислоте;
избыточность (вырожденность) — это значит, что одна ами- нокислота может иметь несколько кодовых триплетов (серин, аланин и др.). Только триптофан и метионин имеют по одно- му кодону (табл. 3);
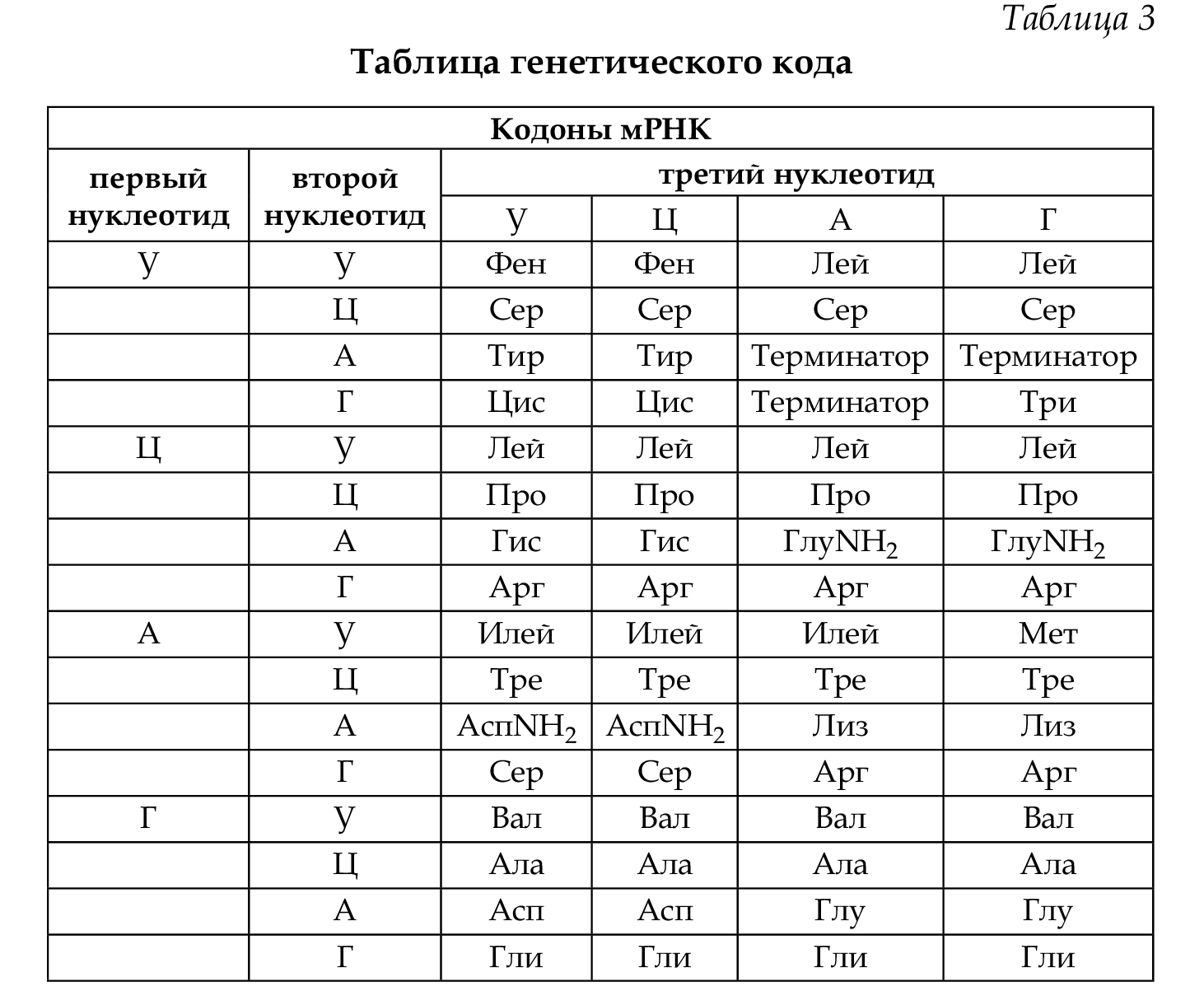
Неперекрываемость — это значит, что один нуклеотид может входить в состав только одного триплета;
Однозначность — это значит, что один кодон соответствует только одной аминокислоте.
Оба этапа относятся к матричным процессам и требуют нали- чия молекулы-матрицы, специфического фермента, энергии и осу- ществляются по правилу комплементарности. Участок молекулы ДНК, структурно-функциональный эквива- лент гена, на котором происходят транскрипционные процессы, на- зывается цистрон (см. рис. 39); он состоит из последовательностей, соответствующих структурному гену, и регуляторных последова- тельностей: промотора и терминатора (трейлера).
Промотор — специализированный участок (сайт) молекулы ДНК (протяженностью несколько десятков нуклеотидов), расположенный перед структурным геном. Промотор состоит из двух частей: 1. участок связывания РНК-полимеразы (ТАТААТ — ТАТА-бокс); 2. участок связывания регуляторного белка. Между этими двумя сайтами расстояние около 20 пар нуклеотидов.
Терминатор — специализированный участок молекулы ДНК, расположенный непосредственно за кодирующей последовательно- стью. Первая часть терминатора представлена участком, состоящим из GС нуклеотидов (GС богатый участок), а другая — несколькими адениловыми нуклеотидами (полиадениловый участок).
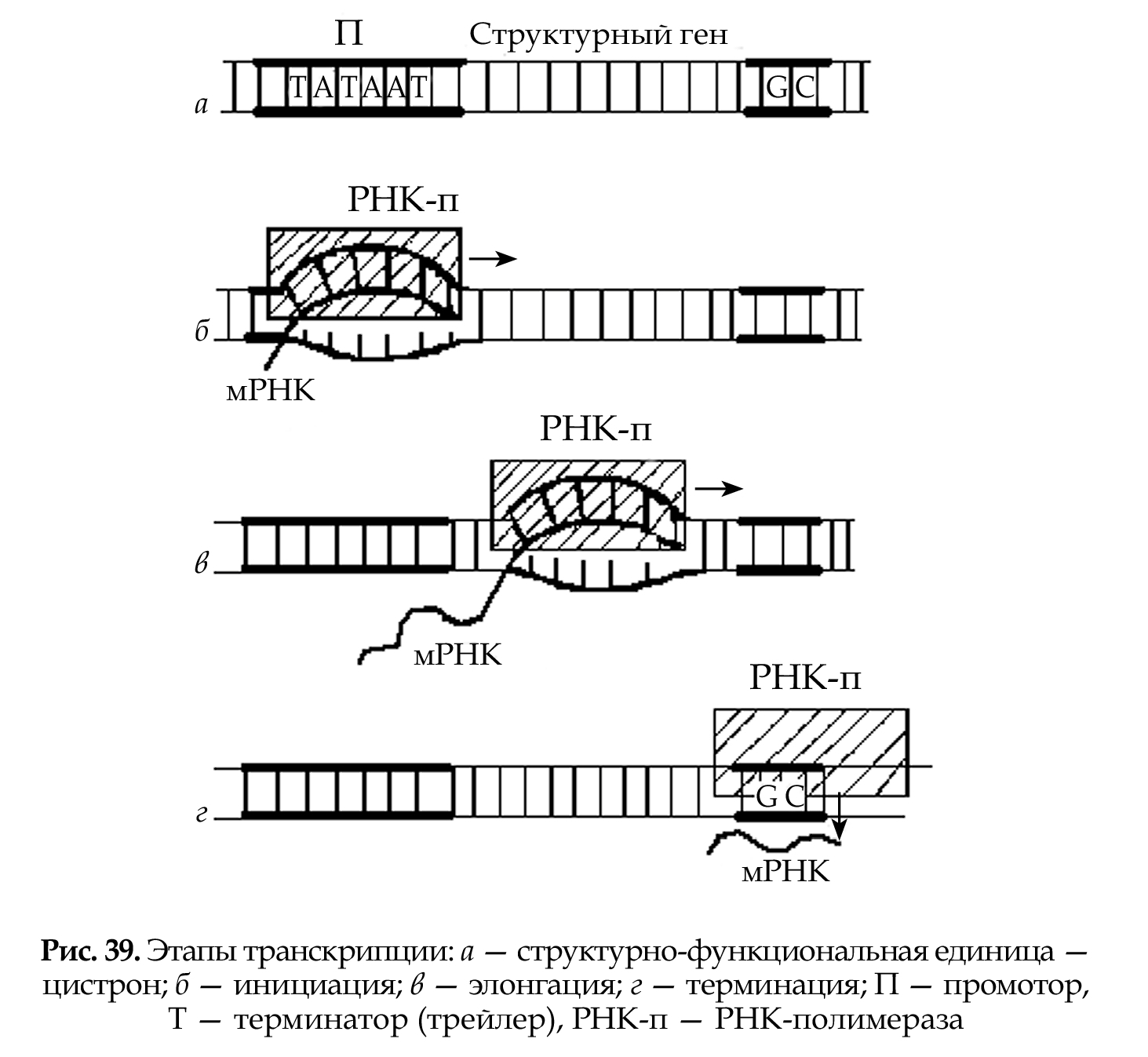
Инициация — начинается со связывания РНК-полимеразы с промотором и началом разъединения цепей ДНК. Синтез мРНК начинается на лидерном участке структурного гена, кото- рый состоит из 3–12 нуклеотидов; начинается с А (у прокариот иногда G) нуклеотида. На этом этапе синтезируемая мРНК не- прочно связана с транскрипционным комплексом (одна цепь ДНК, РНК-полимераза) и легко выходит из него. В этом случае инициация начинается вновь. Затем РНК-полимераза конформируется, связь мРНК с транскрипционным комплексом стабилизируется.
Элонгация — РНК-полимераза ускоряет свое продвижение по ДНК, разъединяя цепи и обеспечивая комплементарное соединение ну- клеотидов. Синтезированная часть молекулы мРНК высвобождается из комплекса, а за РНК-полимеразой восстанавливается двухцепо- чечная структура ДНК.
Терминация — РНК-полимераза проходит участок терминатора, богатый GС парами, а затем присоединяет к себе регуляторный бе- лок, после чего транскрипционный комплекс разрушается, транс- крипция прекращается.
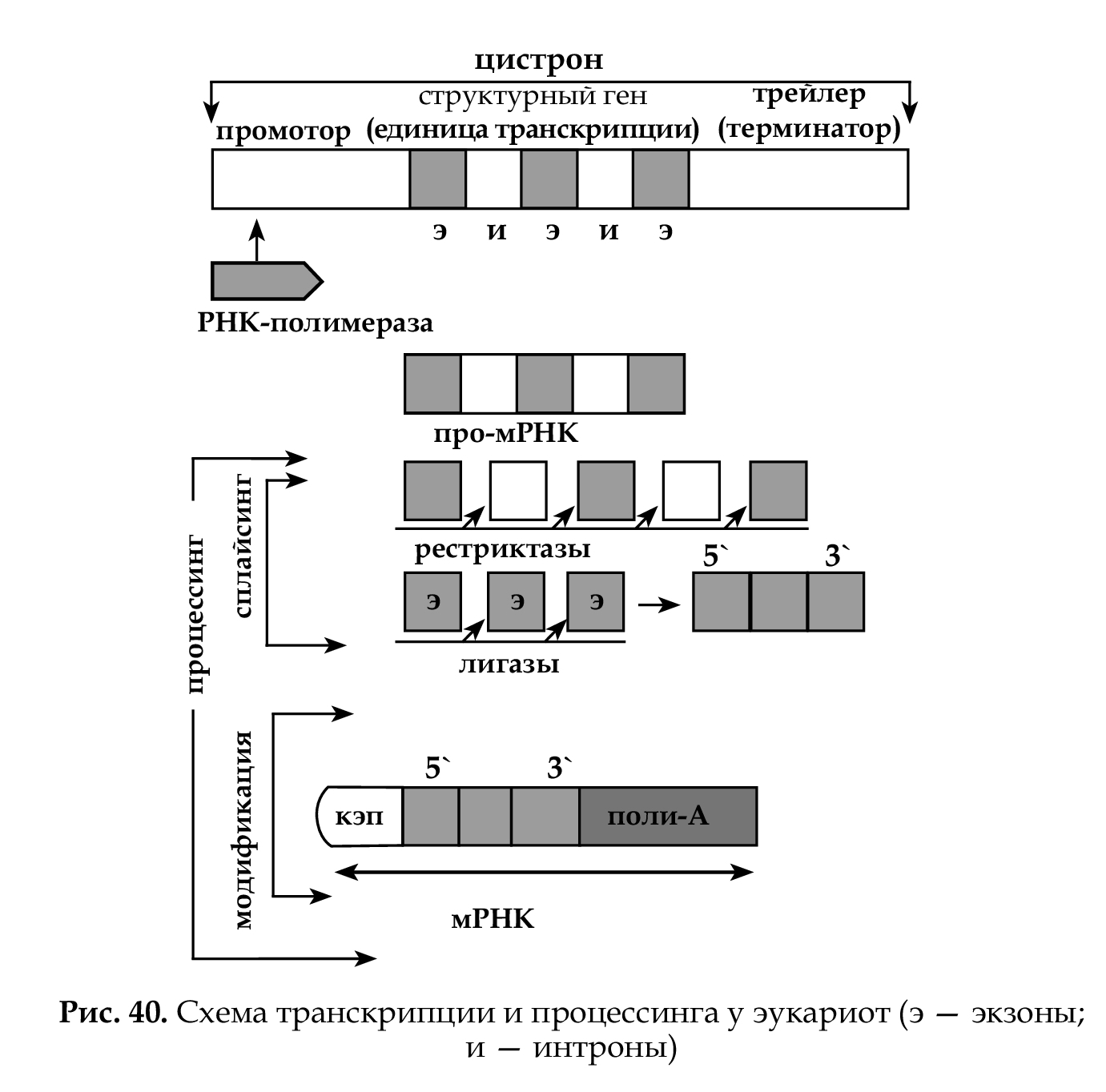
У прокариот синтезируется мРНК, которая может сразу слу- жить матрицей для следующего этапа биосинтеза белка — транс- ляции. У эукариот, в связи с тем, что кодирующие последователь- ности прерываются некодирующими, первичный транскрипт не может служить матрицей при трансляции и еще в кариоплазме подвергается сложному процессу созревания, который называется процессингом , в результате которого проматричная РНК (про- мРНК) превращается в матричную (мРНК) (рис. 40).
Биологический смысл трансляции — перевод информации с по- следовательности нуклеотидов мРНК на последовательность амино- кислот (расшифровка генетического кода) в процессе образования полипептидной цепи (первичной структуры белковой молекулы). Трансляция протекает на рибосомах, расположенных в гиало- плазме, или на мембранах ЭПС. В этом процессе принимают участие все виды РНК: мРНК, рРНК, тРНК. Молекула мРНК связывается с рибосомой; молекулы рРНК входят в состав субъединиц рибосом и выполняют определенные функции; тРНК транспортирует ами- нокислоты из гиалоплазмы к рибосомам. В состав субъединиц рибосом входят разнообразные рРНК, разли- чающиеся по константе седиментации (единица Сведберга — S). Так, в состав малой субъединицы у прокариот входит рРНК 16S, а у эукариот — рРНК 18S, а в состав большой субъединицы у прокариот входят рРНК 5S и 23S, ау эукариот 5S, 5,8S и 28S. Кроме того, в состав рибосом входят разнообразные белки (у прокариот — более 50 уникальных молекул, у эукариот — более 70).
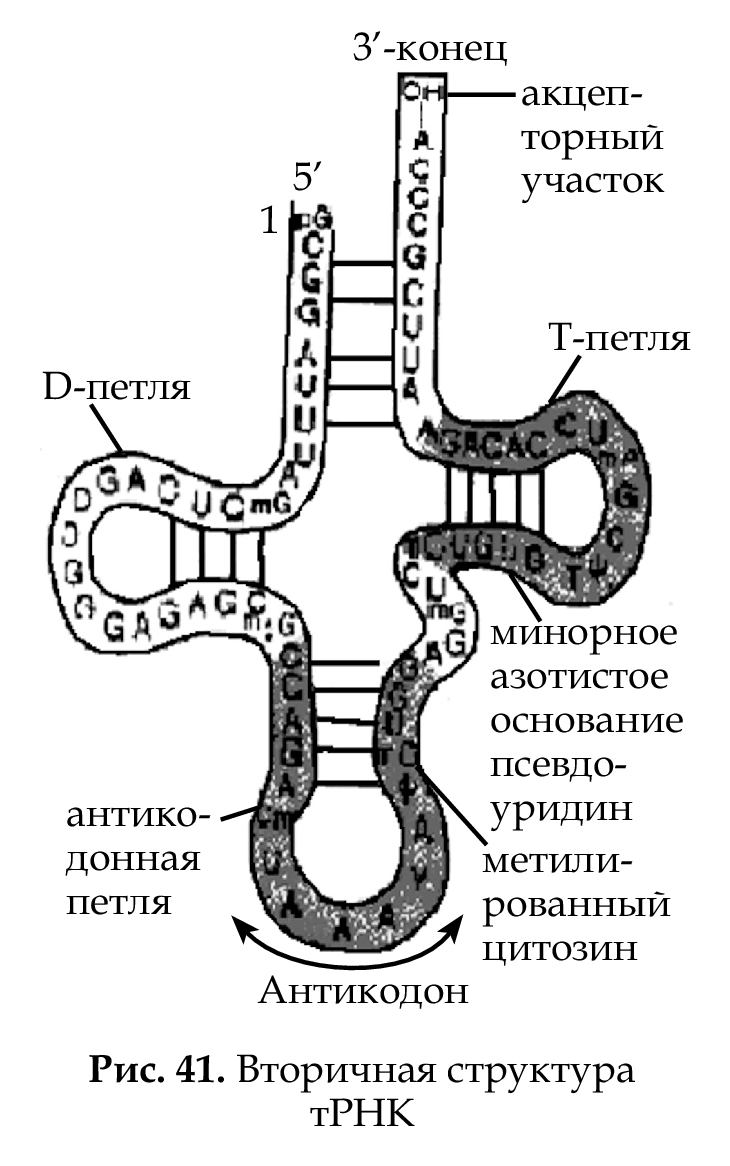
Набор рРНК и белков определяет константу седиментации (S) малой и большой субъединиц рибосом (у прокариот 30S и 50S, у эукариот 40S и 60S) и об- щую константу седиментации (S) рибосом (у прокариот 70S, у эукариот 80S). Молекулы тРНК (рис. 41) — сравнительно короткие молеку- лы, состоящие из 71–94 нуклеотидов. 5'-конец обычно начинается с гуанилового нуклеотида (G), фосфорилирован; 3-'конец (акцепторный участок) всегда заканчивается последовательностью CCA (ЦЦА), соединен с группой −OH или с аминокислотой. В со- став тРНК входят несколько необычных азотистых оснований (минорных): инозин (Y), псевдоуридин (Ψ), дигидроуридин (D); часто встречаются метилированные обычные азотистые основания. Внутримолекулярные комплементарные связи приводят к обра- зованию конфигурации молекулы «клеверный лист». Сдвоенные участки — ветви (стебли), а одноцепочечные участки — петли. Та- ким образом, формируются 4 ветви и 3 петли. Ветви: акцепторная, D (включает основание D), антикодоновая, T (включает псевдоуридин). Петли: D, Т и антикодоновая.
В клетке содержится более 60 разных тРНК. Узнавание тРНК своей аминокислоты называется рекогниция, а связывание с ней — акцеп- ция. Те тРНК, которые способны акцептировать одну и ту же амино- кислоту, называются изоакцепторными. Трансляция также состоит из трех этапов: инициация, элонгация, терминация (рис. 42 ,на примере прокариот).
Инициация.
Начало матричного синтеза (трансляции) происходит поэтапно:
1. связывание мРНК с малой (30 \S) субъединицей рибосомы;
2. установка в пептидильном центре (Р) инициирующего кодона АУГ (AUG);
3. связывание тРНК–формил-метионин (тРНК–f-met) с кодоном АУГ (образование инициирующего комплекса);
4. присоединение большой (50S) субъединицы рибосомы;
5. образование комплекса кодон–антикодон в аминоацильном (А) центре и образование пептидной связи между формил-метиони- ном и второй аминокислотой (образование дипептида);
6. транспозиция рибосомы (перемещение) по мРНК на один три- плет (при этом первая тРНК покидает рибосому, вторая тРНК,
с которой связан дипептид, перемещается из А- в Р-центр, а в А-центре появляется новый кодон).
Элонгация.
Углубление и ускорение процесса трансляции, результатом чего является наращивание полипептидной цепи. Состоит из многократ- но повторяющихся этапов:
1. транспортировка аминокислот в рибосому с помощью тРНК;
2. образование комплекса кодон–антикодон в А-центре и образо- вание пептидной связи между аминокислотами;
3. транспозиция рибосомы по мРНК на один триплет.
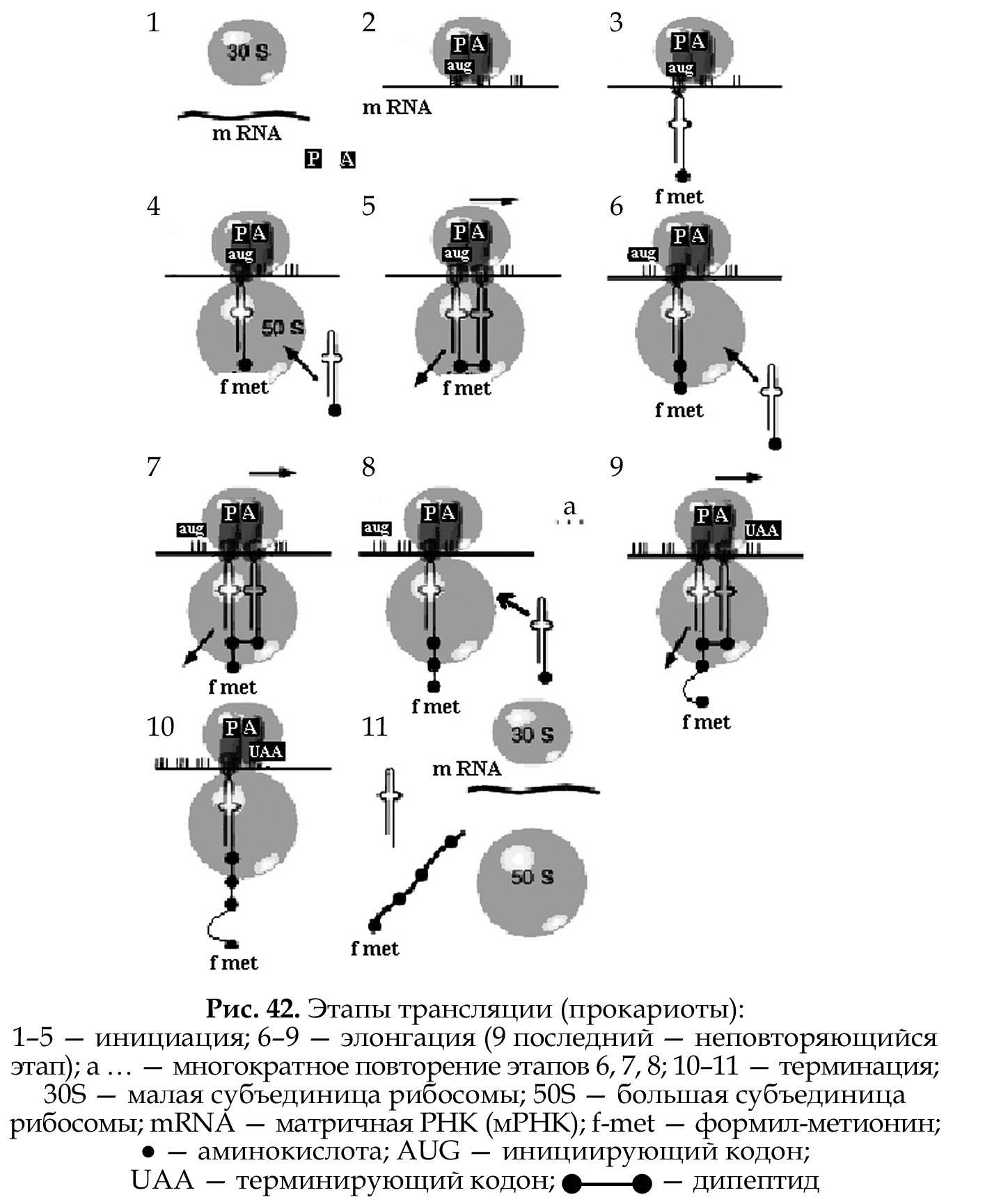
Терминация
Окончание трансляции:
1. появление в А-центре после очередной транспозиции рибосо- мы терминирующего кодона (УАА, УАГ, УГА);
3. в P-центре дестабилизируется и утрачивается связь между тРНК и мРНК;
4. полипептид отщепляется от тРНК;
5. мРНК покидает рибосому;
6. рибосома диссоциируется на субъединицы.
Самовоспроизведение ДНК (процесс, приводящий к удвоению молекул ДНК), называется репликацией. Репликация необходима пе- ред делением клетки, чтобы каждая дочерняя клетка обладала таким же объемом информации, как и материнская. Репликация — реак- ция матричного синтеза, матрицей является материнская ДНК, про- цесс протекает по правилу комплементарности, поэтому дочерние молекулы идентичны материнской. В каждой новой молекуле одна цепь — старая (материнская), которая служила матрицей, а вторая — вновь синтезированная, дочерняя (полуконсервативный способ). Таким образом, биологическое значение репликации ДНК — струк- турная и функциональная преемственность клеток (сохранение специфичности клеток) при воспроизведении себе подобных.
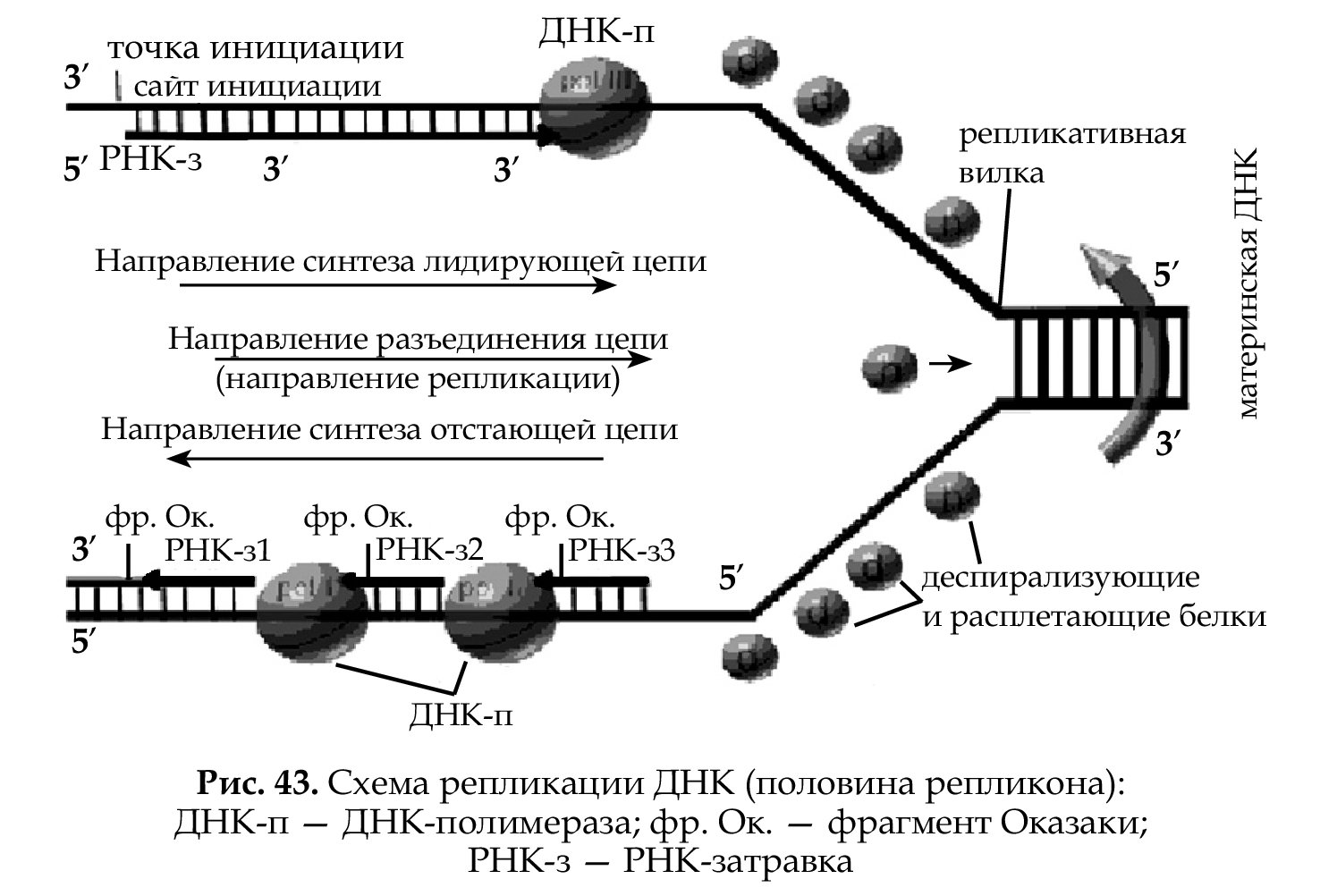
Участок молекулы ДНК, на котором идет репликация, называется репликон. У прокариот репликон один, а у эукариот на одной моле- куле ДНК может быть несколько репликонов. В области репликона цепи разъединены; участки, ограничивающие репликон— реплика- тивные вилки (рис. 43).
Процесс репликации идет по-разному на противоположных це- пях, а также вправо и влево от точки инициации на одной цепи. Это определяется несколькими моментами:
1. Направление разъединения цепей одно для обеих цепей.
2. Цепи ДНК антипараллельны.
3. Разъединение цепей идет в обе стороны от точки инициации (середины репликона).
4. Фермент ДНК-полимераза работает только в одном направле- нии, соединяя нуклеотиды от 5-го положения последующего нуклеотида к 3-му предыдущего.
Иногда в клетке проходит многократная репликация ДНК без по- следующего деления клетки и возникают «многонитчатые» гигантские хромосомы, содержащие сотни и тысячи молекул ДНК. Такие хромо- сомы называются политенными . Биологический смысл политении — резкое увеличение количества определенных генов и, как следствие, резкая интенсификация синтеза определенного белка (см. рис. 49).







