Современное состояние эволюционной теории
На современном этапе развития эволюционного учения различают два уровня эволюционных процессов - микроэволюцию и макроэволюцию.
1. Микроэволюция - эволюционные изменения, происходящие на поиуля- ционном уровне и завершающиеся образованием новых видов. Элементарной единицей эволюции является популяция. Популяция - минимальная самовоспроизводящаяся группа особей одного вида, свободно скрещивающихся между собой, длительно существующая на определенной территории, образующая самостоятельную генетическую систему. Любая популяция имеет морфофизиологическую, экологическую и генетическую характеристику.
В популяции действуют элементарные эволюционные факторы: мутационный процесс, популяционные волны, изоляция, дрейф генов, естественный отбор. Взаимодействуя, они могут приводить к изменению генофонда популяции (элементарное эволюционное явление). Мутационный процесс (спонтанный мутагенез) поставляет новые аллели (элементарный эволюционный материал), которые разнообразят генетический состав популяции (обогащают генофонд).
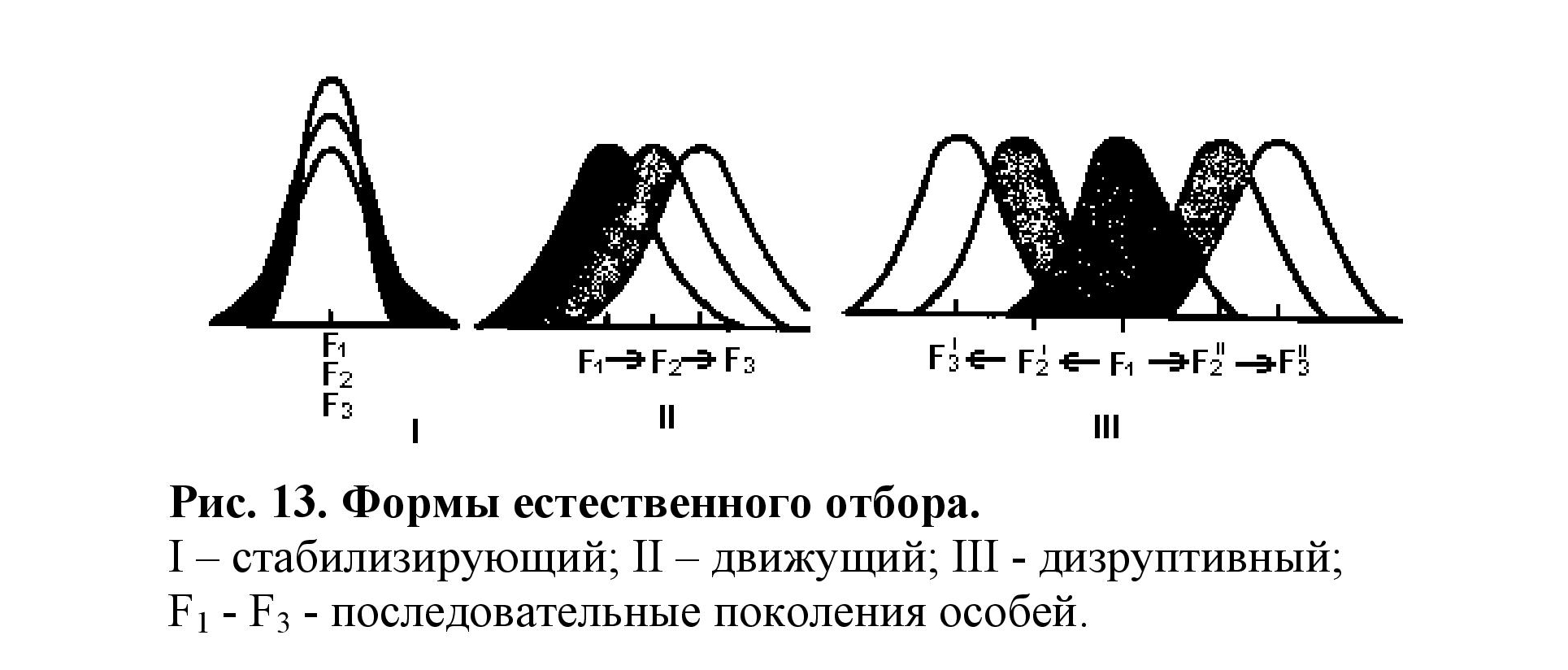
Популяционные волны - периодические и непериодические резкие колебания численности особей в популяции. Изоляция- возникновение любых барьеров (пространственных д биологических), ограничивающих панмиксию. Естественный отбор - постоянно протекающий процесс, обеспечивающий избирательную выживаемость и размножение одних особей (наиболее приспо-собленных) и гибель других. Основные формы естественного отбора (рис. 13):
• стабилизирующий - направлен на поддержание в популяции среднего, ранее сложившегося значения признака или свойства. (Пример: после снегопада и сильных ветров выживают птицы со средними - «нормальными» крыльями).
• движущий - ведет к последовательному изменению фенотипа, определяет сдвиг средних значений признаков. (Пример: индустриальный мела- низм берёзовой пяденицы).
• дизруптивный - разрывает популяцию на несколько групп, определяет развитие признака в разных направлениях. (Пример; появление на океанических островах насекомых бескрылых и с более длинными крыльями по сравнению с материковыми).
Естественный отбор определяет целесообразность изменений для конкретных условий существования.Эволюционные процессы в популяциях завершаются образованием подвидов, а затем видов. Видообразование - процесс возникновения новых видов путем преобразования существующих. Пути видообразования:
• симпатрическое - видообразование осуществляется на одной территории, новый вид возникает внутри ареала исходного. (Пример: полиплоидные формы, раннецветущие и позднецветущие формы лютика).
• аллопатрическое видообразование - основано на пространственной изоляции. (Пример: разные виды соболя - европейский, сибирский).
Способы видообразования различны, (рис. 14) Основными являются:
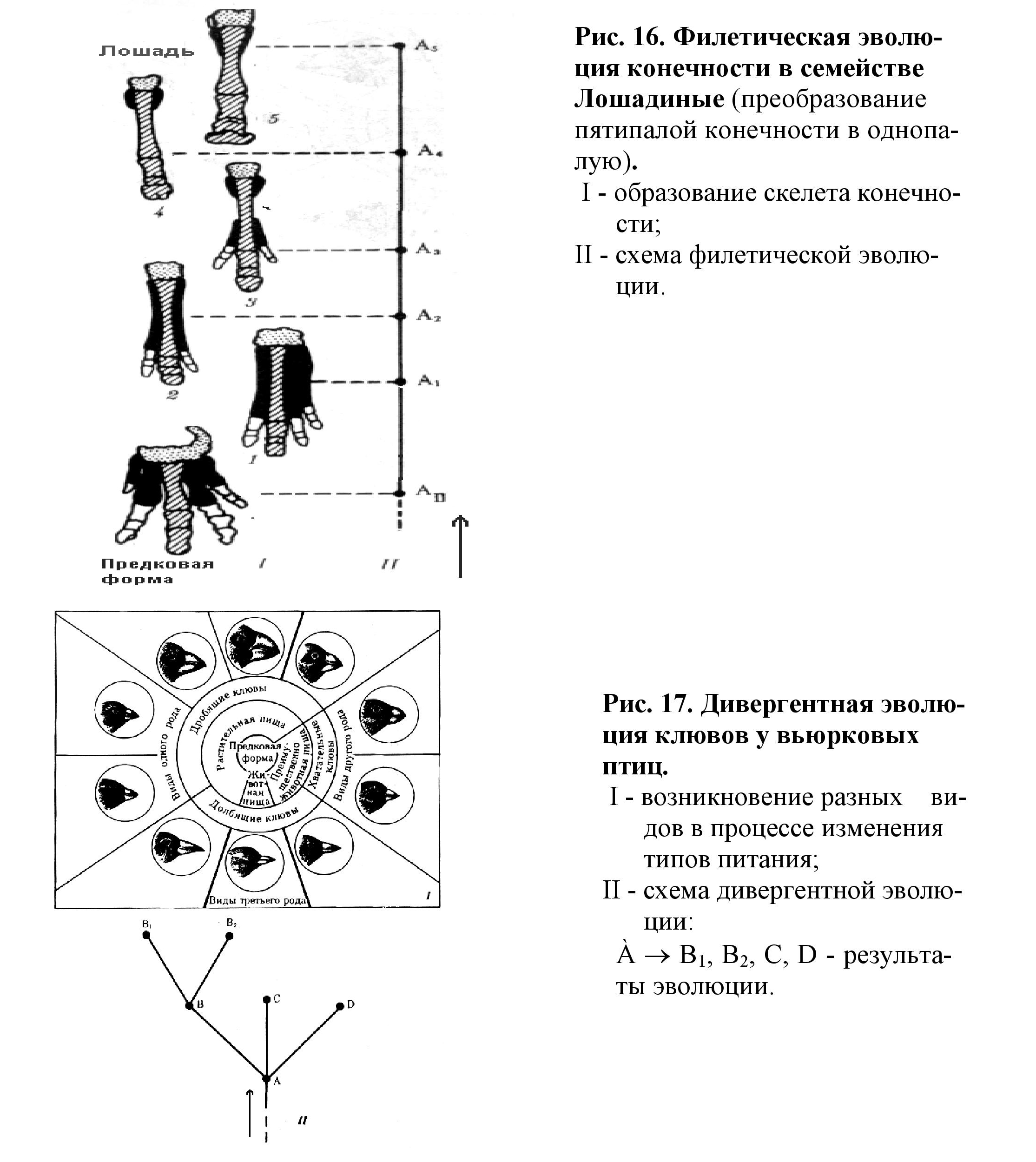
• филетическое - постепенное превращение во времени одного вида в другой. (Пример: развитие предков лошадей по прямой линии).
• дивергентное - разделение первоначально единого вида на два или более новых. (Пример: разнообразие вьюрков на Галапогосских островах).
• гибридогенное - путем гибридизации. (Пример: растение рябинокизильник в Сибири).
2. Макроэволюция - процесс формирования таксонов надвидового ранга. Процесс можно изучать методами палеонтологии, сравнительной анатомии и сравнительной эмбриологии, молекулярной биологии и генетики. В основе макроэволюции лежат закономерности наследственности и изменчивости, раз-
витие приспособлений и естественный отбор, который является направляющей
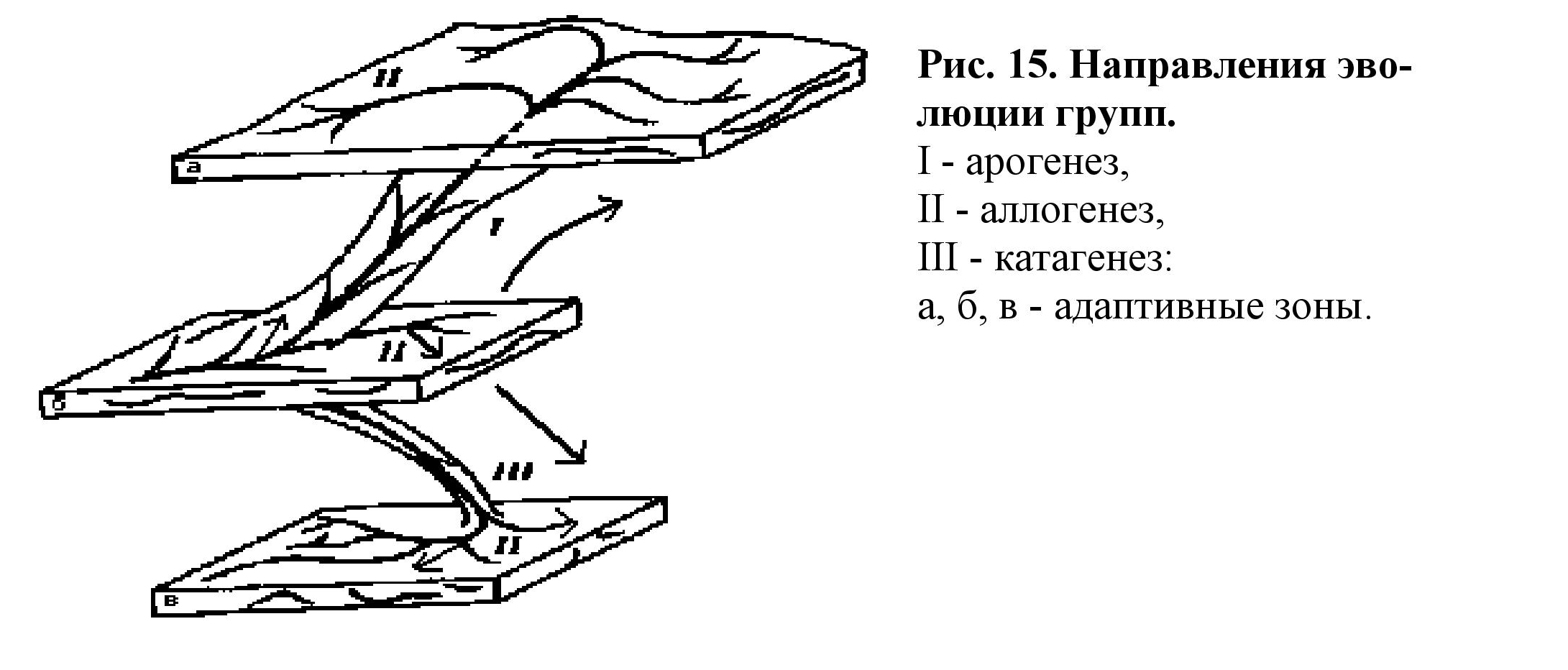
и контролирующей силой. Главные направления эволюции групп (рис. 15):
• Арогенез - развитие группы с расширением адаптивной зоны и с
освоением новой среды обитания. Осуществляется на основе ароморфозов (на-
пример, развитие земноводных).
• Аллогенез - развитие группы в пределах одной адаптивной зоны
осуществляется на основе идиоадаптаций (например, появление разнообразных
форм в отряде насекомоядных млекопитающих - землеройка, крот, выхухоль).
• Катагенез - развитие группы с сужением адаптивной зоны (напри-
мер: развитие личиночнохордовых).
Пути эволюции: ароморфоз, идиоадаптация (алломорфоз) и дегенерация. Ароморфоз - морфофизиологический прогресс, это крупные прогрессивные изменения, повышающие организацию и жизнедеятельность организмов и приводящие к арогенезу (например: образование хорды у хордовых, черепа у позвоночных, четырехкамерного сердца у птиц). Идиоадаптация - изменения, определяющие специализацию в данной адаптивной зоне, приводят к адаптивной радиации (например, разные формы клюва у птиц). Дегенерация - морфофизиологический регресс - это изменения, приводящие к упрощению организации и занятию новой адаптивной зоны (например, редукция пищеварительной системы у ленточных червей). Формы эволюции групп:
1. Филетическая эволюция - это изменения, происходящие в одном филоге-нетическом стволе, приводящие к преобразованию исходного вида (рис. 16).
2. Дивергентная эволюция - расхождение признаков в рамках популяции, вида, возникающее в условиях изменяющейся среды; приводит к возникновению новых видов из одного исходного (рис. 17).
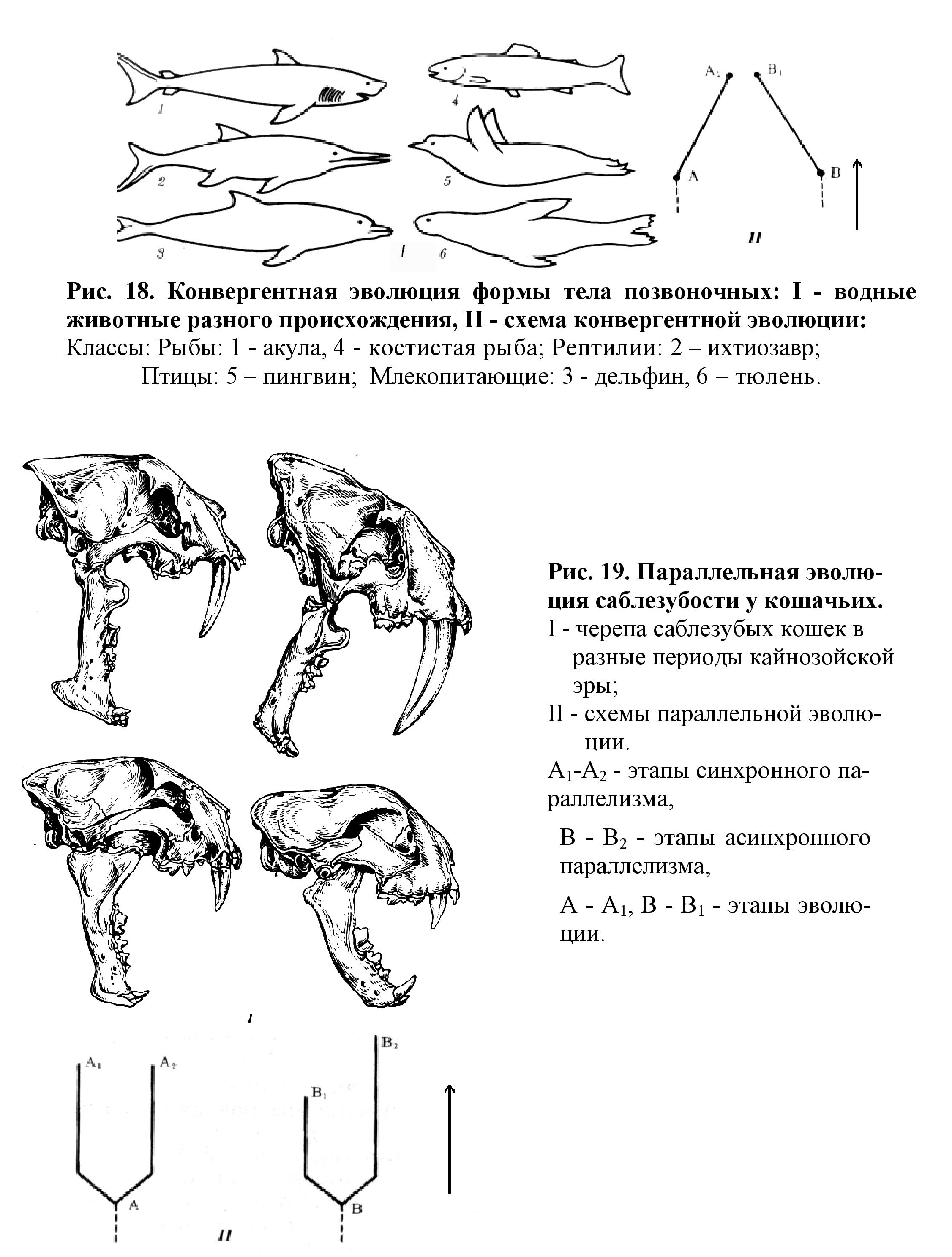
3. Конвергентная эволюция: - формирование фенотипического сходства у неродственных видов, возникающее в сходных условиях существования (обитания и жизнедеятельности) (рис. 18).
4. Параллелизм - формирование сходств у первоначально дивергировавших генетически родственных групп, происходящее в сходных условиях существования (рис. 19).
5. Сим биогенез - происхождение организмов путем соединения двух или нескольких существ, вступающих в симбиоз (лишайник).
Генетика популяций
Генетическая характеристика популяций определяется генофондом. Гено-
фонд - это совокупность всех аллелей особей популяции. Генофонд определя-
ется генетической гетерогенностью. Генетическая гетерогенность - наличие в популяции нескольких аллель-
ных вариантов гена (минимум двух). Чем большее число генов имеют аллель-
ные варианты, тем выше генетическая гетерогенность популяции. Она поддер-
живается мутационным процессом. Наследственный полиморфизм - наличие в популяции на протяжении
многих поколений нескольких генетических форм (генотипов). Он определяет-
ся генетической гетерогенностью и процессом рекомбинации. Различают две
формы наследственного полиморфизма: а) экологический (адаптационный), б)
гетерозиготный, который устанавливается в результате давления положитель-
ного отбора гетерозигот.
Генетически полиморфная популяция в каждый момент имеет приспособ-
ленность и жизнеспособность ниже уровня жизнеспособности популяции, со-
стоящей только из «удачных» генотипов. Различие в степени жизнеспособности
этих двух популяций называется генетическим грузом. Бремя генетического
груза можно оценить, введя понятие - летальный эквивалент. Один летальный эквивалент - совокупное действие неблагоприятных алле-
лей равное действию одного летального гомозиготного состояния, приводящего
организм к гибели до наступления репродуктивного периода.
Генетические характеристики по-
пуляции можно изучать с помощью за-
кона Харди-Вайнберга, который позво-
ляет вычислить частоты аллелей и соот-
ношение генотипов в популяции; эти по-
казатели остаются постоянными, если ге-
нофонд популяции не изменяется.
Если доминантный аллель А обо-
значить как р, а рецессивный аллель а
как q, то в потомстве свободно скрещи-
вающихся особей должны быть сле-
дующие генотипы (табл. №9). Сумма рецессивных и доминантных аллелей
одного гена в популяциях равна 1, т.е. p+q =1. Зная частоту встречаемости q,
можно вычислить частоту встречаемости р.
Допустим, q = 0,3, следовательно, р = 1 - 0,3 = 0,7. Подставив эти значения в формулу, можно рассчитать количество разных генотипов в популяции (см. задачу на стр. 47 ).
Закон Харди-Вайнберга справедлив для идеальной популяции, которая характеризуется следующими особенностями:
• бесконечно большая численность
• панмиксия (свободное скрещивание)
• отсутствие мутаций (отсутствие возникновения новых аллелей в популяции)
• отсутствие миграции особей из других популяций (отсутствие привнесения новых аллелей извне)
• отсутствие отбора
Реальные популяции отличаются от идеальной по всем параметрам, однако, закон Харди-Вайнберга можно применять (с некоторыми допущениями) для определения генотипического состава популяции по определенным аллелям, в том числе, и в популяциях человека (см. задачу па стр. 40).
В небольших популяциях статистические закономерности Харди- Вайнберга же соблюдаются, в них начинают действовать генетико- автоматические процессы (дрейф генов). Суть этих процессов в том, что постепенно из популяции исчезают гетерозиготы, и вся популяция становится гомозиготной (АА или аа). Таким образом, из популяции исчезает один аллель определенного гена, генетическая гетерогенность популяции снижается, генофонд обедняется. Чем малочисленнее популяция, тем более интенсивно протекают геиетико-автоматические процессы. Дрейфу генов способствует также изоляция.
Особенности действия эволюционных факторов в популяциях человека
В антропогенетике популяцией называют группу людей, проживающих на определенной территории, представители которой могут свободно вступать в брак.
Демографическими показателями популяции людей являются численность особей, уровень рождаемости и смертности, возрастной и половой состав, экономическое состояние, уклад жизни. Популяции человека делятся на крупные, малые (субпоцущщйи) :шш демы и изоляты.
Крупные популяции - численность более 4000 человек расселены на зна-чительных территориях (не урбанизированные популяции). В таких популяциях распределение частот аллелей и генотипов подчиняется закону Харди- Вайнберга (табл. №9).
Демы - популяции численностью примерно 1500 4000 человек.
Изоляты - самые маленькие популяции - не более 1500 человек.
Генетически человеческие иопуляции характеризуются генофондом, который во многом зависит от размеров группы и структуры браков. Генетическая гетерогенность человеческих популяций значительна. У человека гетеро- зиготность составляет около 7%, при расчете на 100000 структурных генов разнообразие гамет составит 2 . Отсюда и высокая степень наследственного полиморфизма. Летальный эквивалент человека равен 3 - 5.
С возникновением человека как социального существа биологические факторы эволюции постепенно ослабляют свое действие, и ведущее значение в развитии человечества приобретают социальные факторы. Однако Человек как биологический вид подчиняется законам, действующим в живой природе.
Мутационный процесс — эволюционный фактор, который сохраняет свое значение в человеческом обществе. Его действие сходно с таковым у других организмов но средней частоте мутирования, по генетико-физиолошческим характеристикам. Па человека на начальных этапах эволюции преимуществен^ пое действие оказывали факторы спонтанного мутагенеза. В настоящее время давление мутационного процесса на генофонд человечества усиливается в результате действия индуцированиого мутагенеза.
Изоляция. Специфическими для человеческого общества являются соци-альные формы изоляции, зависящие от разнообразия культур, экономических укладов, религаозных и морально-этических установок. Высокая степень репродуктивной изоляции малочисленных человеческих популяций на протяжении многих поколений создавала условия для дрейфа генов. Примером дрейфа генов служит эффект родоначальника. Так, члены секты амишей штата Пенсильвания произошли от трех супружеских пар. В этом изоляте обнаружено 55 случаев сочетания карликовости с многопалостью, в то время как в мире всего зарегистрировано только 50 случаев этой аномалии.
Популяционные волны в настоящее время не имеют существенного эволюционного значения.
Естественный отбор. Смена биологических факторов развития социальными привела к тому, что в человеческих популяциях отбор утратил функцию видообразования. Он действует, в основном, во время внутриутробного развития,, выражается в таких формах, как несостоявшаяся беременность, спонтанный аборт, мертворождение, детская смертность, стерильность и имеет стабилизирующий характер.
Пример решения задачи на закон Харди-Вайнберга
Задача. В странах Скандинавии фенилкетонурия (аутосомно-рецессивное наследование) встречается с частотой 1:40000. Рассчитайте долю гетерозиготных носителей рецессивного аллеля.
Решение. Исходя из закона Харди-Вайнберга, 3 возможных генотипа АА, Аа и аа распределяются с частотой р2, 2pq и q2 соответственно. Генотип больного - аа, следовательно, q2=l/40000, q=l/200. Частота доминантного аллеля: р=1 -q=l -1/200=199/200.
Частота гетерозигот= 2pq=2xl99/200xl/200=2xl99/40000=0,00995=0,01 (1 %).
При найденных частотах аллелей в популяции численностью 40000 человек имеется один больной фенилкетонурией (аа) и 400 носителей (Аа). Остальные члены гомозиготны по благоприятному доминантному аллелю (АА).