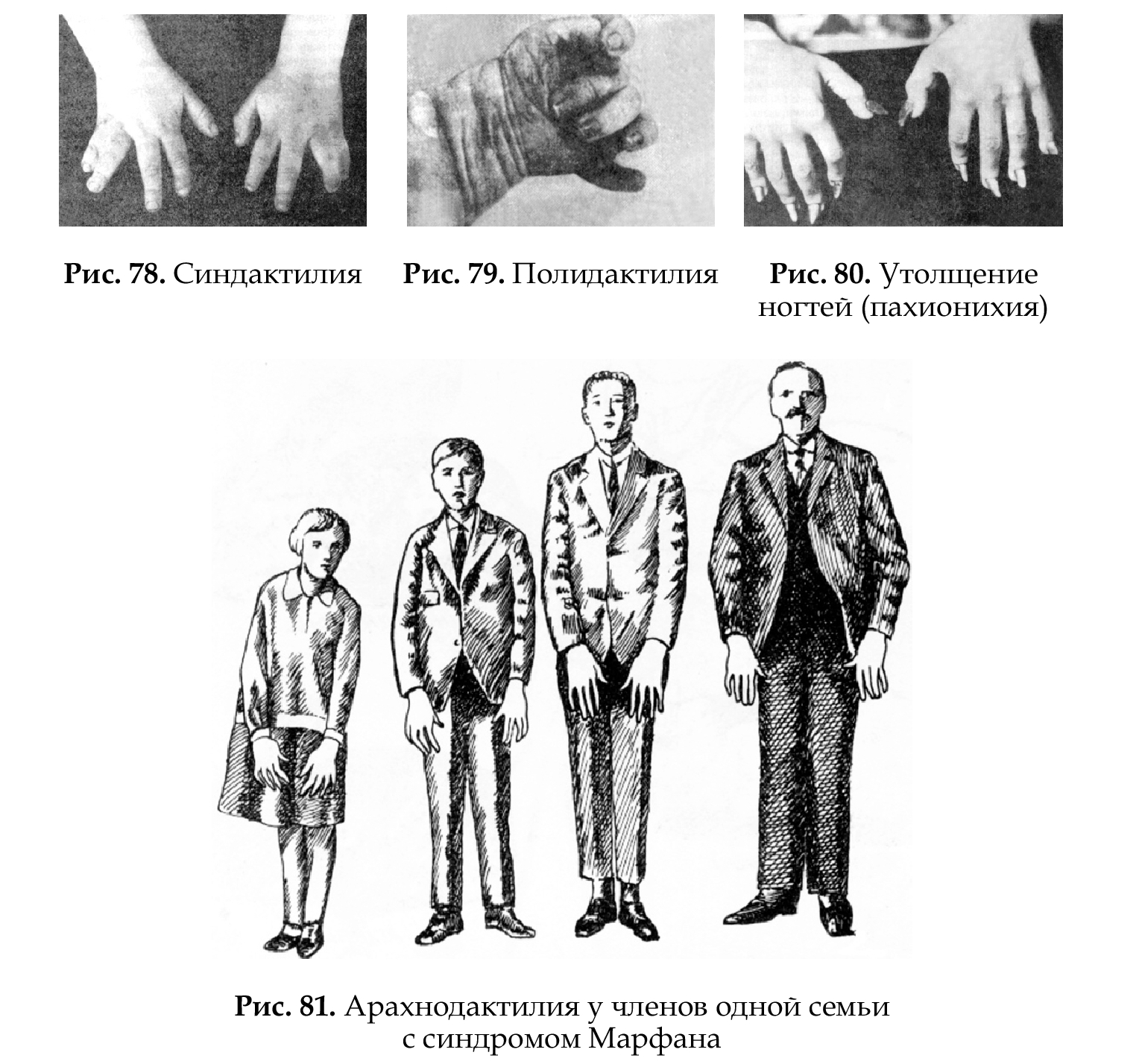
Наследственные болезни — болезни, связанные с качественным и количественным нарушением наследственного материала. Существуют разнообразные классификации наследственных болезней, которые широко используются в клинике и в теоретических исследованиях; основные из них:
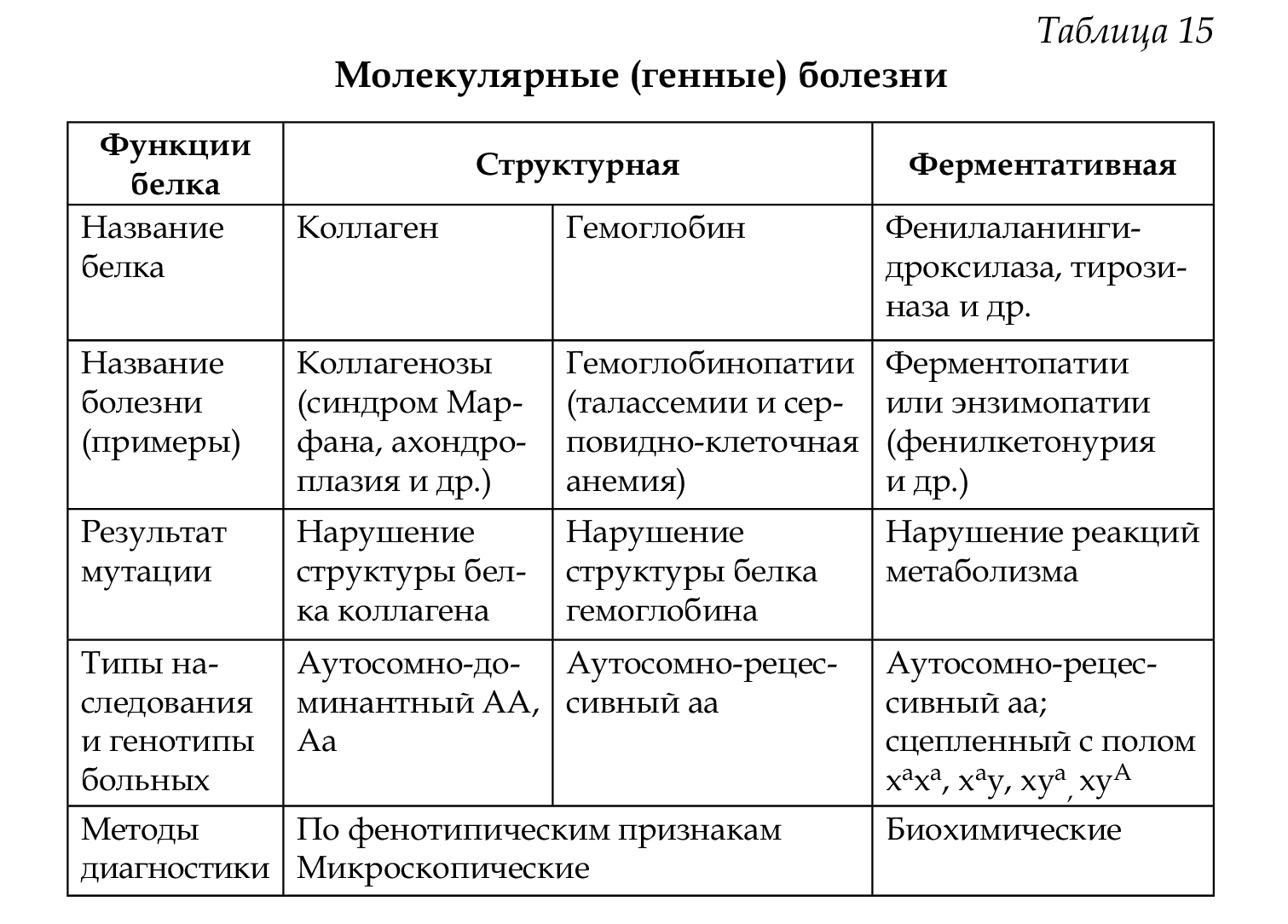
• по уровню нарушения генетического материала: молекулярные (генные) болезни (табл. 15) и хромосомные;
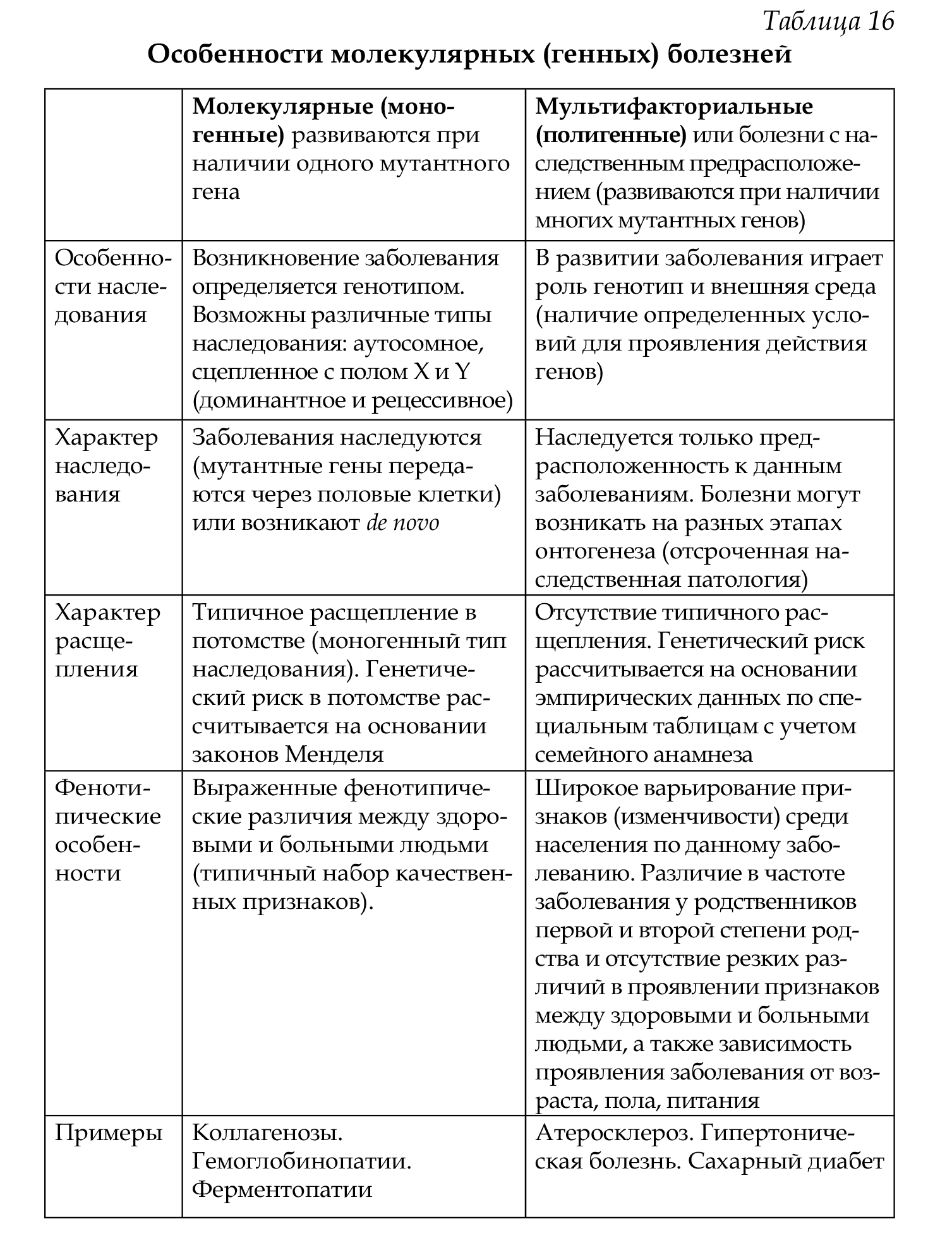
• по способу наследования признака: моногенные и полигенные (мультифакториальные) (табл. 16).
Наследственные болезни наследуются не все, нельзя дословно воспринимать это название, часть из них появляется de novo.
10.2 Молекулярные (генные) болезни
Причина возникновения — генные мутации (см. соответствующую тему). Механизм развития заболевания: изменение нуклеотидной последовательности ДНК —> изменение мРНК —> изменение белка (структурного или белка-фермента) —> появление патологических признаков —> болезнь.
Энзимопатии могут возникать при нарушении всех видов обмена (см. словарь терминов):
• углеводного — галактоземия, фруктозурия, полисахаридоз, муковисцидозы;
• аминокислотного — фенилкетонурия, алкаптонурия, тирозиноз;
• липидного — болезнь Тея-Сакса, гиперхолестеринемия;
• пуринового и пиримидинового — синдром Леша-Нихана;
• нуклеинового — прогерия;
• минерального — болезнь Вильсона-Коновалова (гепато-церебральная дегенерация), гипофосфатемия (витамин-D- резистентный рахит).
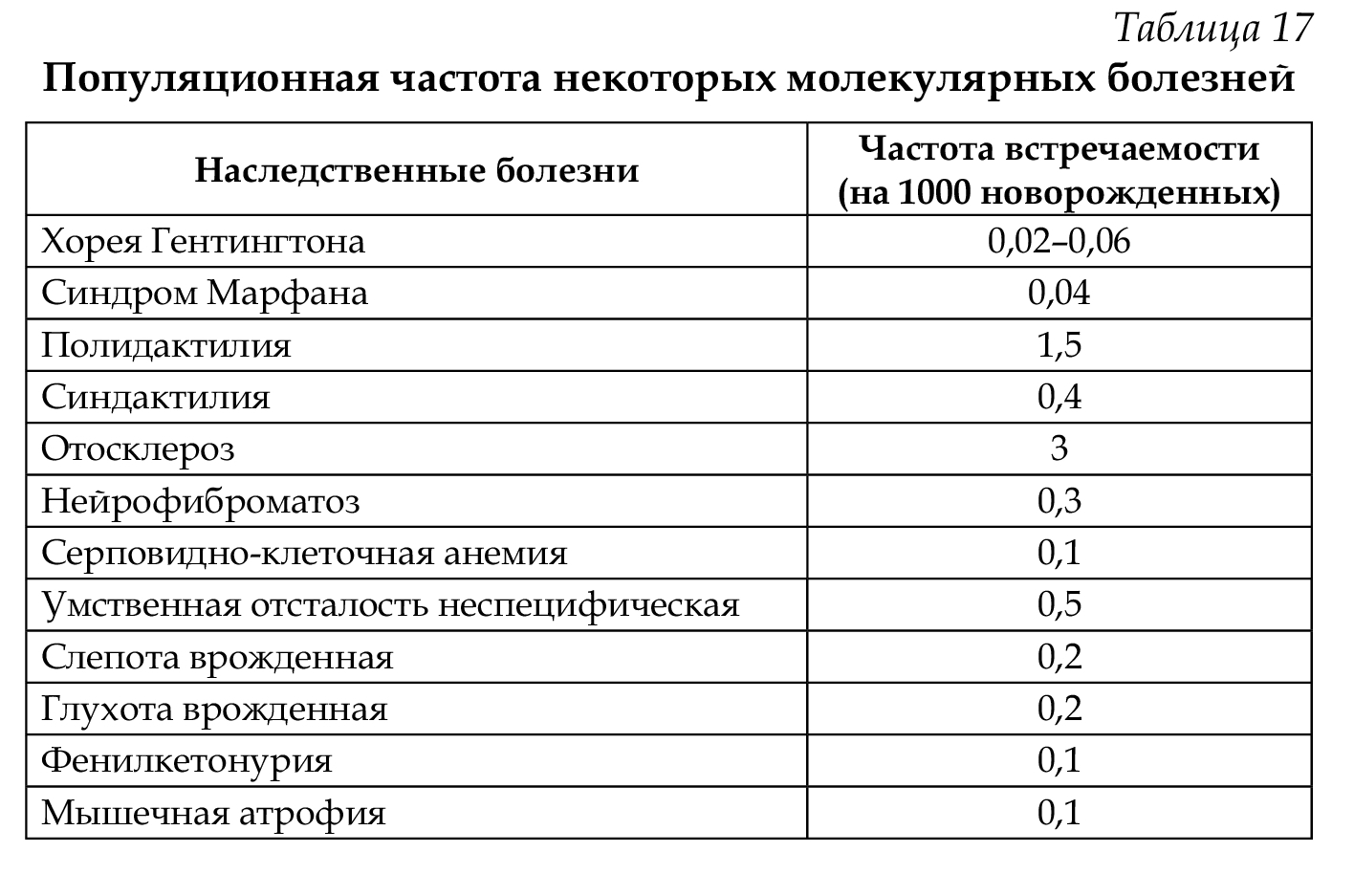
Описаны нарушения обмена гормонов, витаминов, дефекты ферментов эритроцитов и т. д. Молекулярные болезни встречаются в популяциях с различной частотой (табл. 17).
Фенилкетонурия
Моделью для изучения энзимопатий может служить фенилкетонурия. Классическая фенилкетонурия вызвана мутацией гена РАН (пи эй эйч), он картирован (12q 22), идентифицирован и секвенирован (определена последовательность нуклеотидов). Ген РАН относится к мозаичным генам и состоит из 13 экзонов и 12 интронов; он детерминирует синтез фермента фенилаланингидроксилазы — ФАГ.
Заболевание связано с нарушением обмена аминокислоты фенилаланина. В норме аминокислота фенилаланин превращается в аминокислоту тирозин, а тирозин — в пигмент меланин. Мутация гена вызывает уменьшение активности фермента ФАГ, в результате этого фенилаланин не полностью превращается в тирозин. Фенилаланин накапливается в крови и частично превращается в фенилпировиноградную кислоту (ФПК), которая выделяется с мочой и потом — от больных исходит «мышиный запах». ФПК является нейротропным ядом (нарушается формирование миелиновой оболочки вокруг аксонов ЦНС), поэтому у детей развивается повышенная возбудимость, тремор, судорожные эпилептиформные припадки, происходит нарушение высшей нервной деятельности, развивается тяжелая умственная отсталость.
Диагностика фенилкетонурии осуществляется биохимическими (определение фенилаланина в крови и ФПК в моче), микробиологическими (тест Гартри), молекулярно-генетическими и клиническим методами. Предупреждение тяжелых последствий в развитии болезни базируется на диетотерапии: используются белковые гидролизаты с уменьшенным количеством фенилаланина и специаль-ные наборы продуктов (мед, орехи и др.). Лечение детей проводится до 7-10 лет, мозг взрослого человека устойчив к высоким концентрациям ФПК.
При нарушении активности фермента тирозиназы (мутация гена) не происходит превращение тирозина в меланин и возникает альбинизм. У больных наблюдаются слабая пигментация кожи, волос, радужной оболочки, изменения в почках, печени, селезенке.
В ряде случаев развивается алкаптонурия, причиной возникновения которой является генетический дефект фермента оксидазы; в результате этого гомогентизиновая кислота (промежуточный продукт обмена) не расщепляется полностью до воды и углекислого газа, что происходит в норме, а откладывается в соединительной ткани (цвет охры) и выводится с мочой (темная моча).
Данные заболевания наследуются по аутосомно-рецессивному типу, встречаются с относительно высокой частотой 1 : 10000, при частоте мутантного гена у гетерозигот в человеческих популяциях 1 : 50 (1 : 75).
Профилактика: медико-генетическое консультирование на разных этапах онтогенеза, исключение браков между гетерозиготами, а для этого выявление гетерозигот с использованием соответствующих методов. В настоящее время проводится скрининг всех новорожденных на фенилкетонурию, гипотиреоз и муко- висцидоз.
Примеры молекулярных болезней, связанных с изменением структурных белков
Организация и экспрессия генов у прокариот и эукариот
10.4.1. Классификация генов
Элементарной функциональной единицей наследственности является ген, его химическая основа — молекула ДНК. Критерием функции гена является его экспрессия (синонимы: дерепрессия, активация, работа, включение) в процессе реализации генетической информации:
ДНК —> РНК —> белок —> признак
По своему функциональному значению гены классифицируются на структурные, регуляторные и гены-модуляторы. Структурные гены делятся на две группы: гены I, кодирующие структуру белков (полипептидов), гены II — структуру рРНК, тРНК. Регуляторные гены координируют активность структурных генов на уровне клетки и на уровне организма в процессе онтогенеза, а также детерминируют синтез регуляторных белков (ген-регулятор лактозного оперо- на и ген TFM). Наряду с регуляторными генами, есть регуляторные последовательности (промотор, оператор, терминатор, энхансер, элемент перед промотором), функции которых выявляются при вза-имодействии со специфическими регуляторными белками.
Структурные гены делят на гены «домашнего хозяйства», продукты экспрессии которых необходимы постоянно для жизнедеятельности любого типа клеток (гены рРНК, тРНК, гены гистонов, гены тубулинов и др.), и гены «роскоши» — тканеспецифические гены, обеспечивающие специализированные функции клеток, т. е. гены функционально активные только в определенных типах клеток и на определенных стадиях развития организма (гены глобина, ин-сулина, кристаллина, иммуноглобулинов (локус HLA) и др.
Организация генов
Структурные гены различным образом организованы у про- и эукариот. Гены в геноме могут располагаться одиночно (индивидуально от других генов) или группироваться, образуя транскрипционные единицы, опероны (у прокариот) или повторяющиеся гены, кластеры генов (у эукариот).
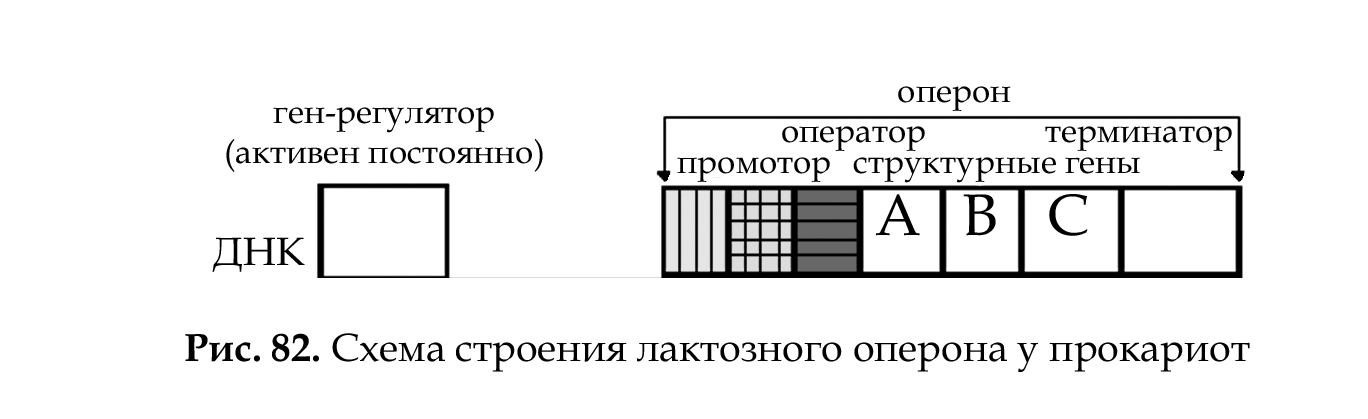
У прокариот (рис. 82) основным типом организации генов являются опероны (например, лактозный оперон кишечной палочки Е. coli).
Оперон Е. coli — это группа структурных генов А, В, С, расположенных друг за другом, которые имеют общий промотор, оператор (нуклеотидные последовательности промотора и оператора перекрываются) и терминатор. Они участвуют в одном метаболическом цикле (в данном случае расщепление лактозы до глюкозы и галактозы) и регулируются координированно. Структурные гены в составе оперона находятся под контролем оператора. Регуляция оперона осуществляется геном-регулятором (см. рис. 84).
У эукариот — основным типом организации генов являются кластеры.
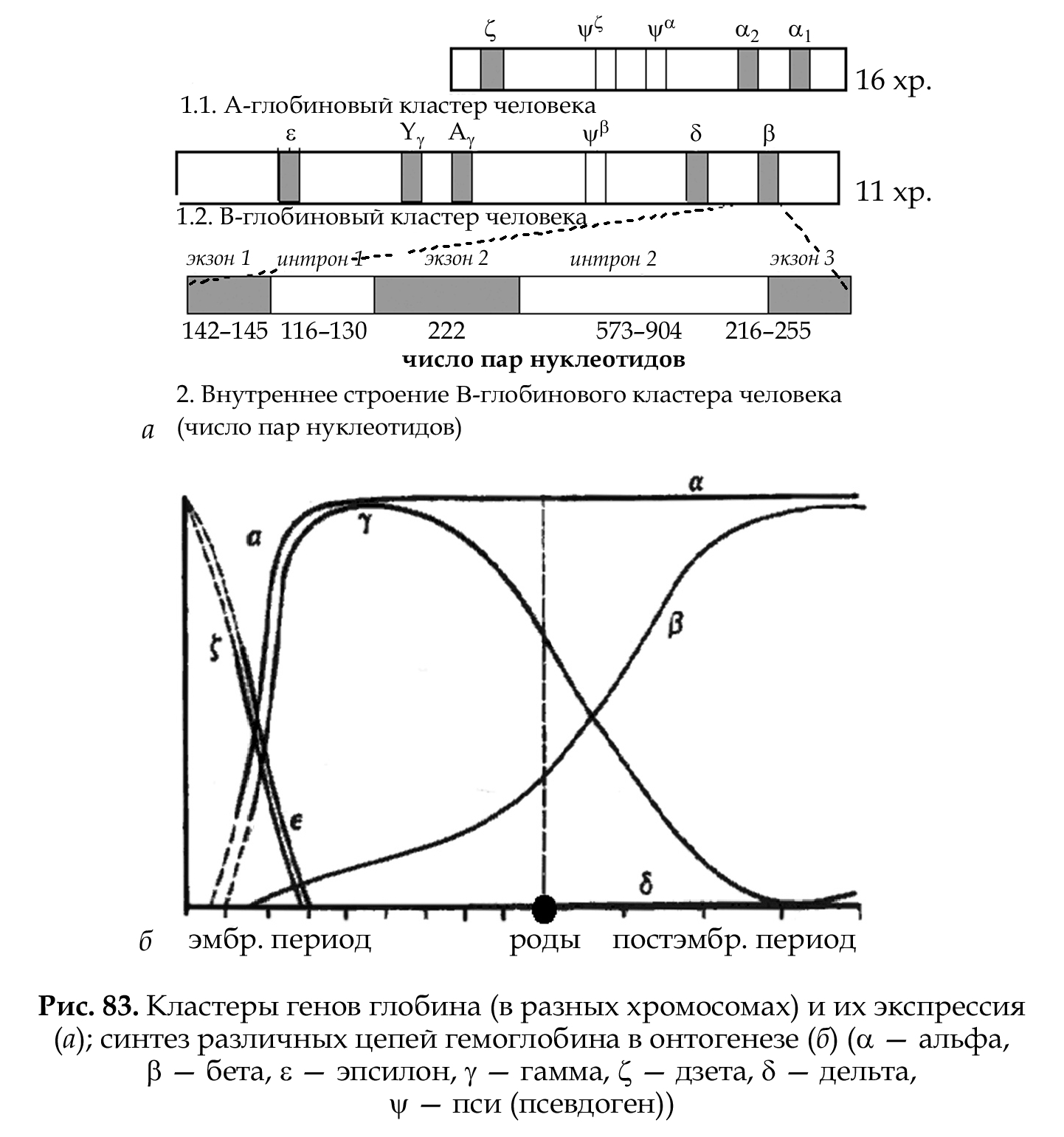
Кластеры генов — это группа родственных генов со сходными функциями, локализованная в определенных участках хромосом. В состав кластера входят активно функционирующие гены и псев- догены (\|/). Часто кластеры являются семейством генов, произошедшим от некоего гена-предка. Классическим примером являются гены глобинов в составе А- и В-кластеров (см. рис. 83). Гемоглобин состоит из гема и белка тетрамера-глобина. Глобиновый тетрамер состоит из двух идентичных P-цепей и двух идентичных a-цепей. Аминокис-лотная последовательность каждой глобиновой цепи кодируется своими собственными генами, входящими соответственно в состав А- и В-кластера. У человека A-кластер располагается в 16 хромосоме, а В-кластер — в 11 хромосоме.
В-кластер занимает участок ДНК размером 50 тысяч пар нуклеотидов и включает в себя функционально активные гены и один псевдоген: ген Е (эпсилон); два гена у (гамма); псевдоген р (бета); ген 5 (дельта) и ген р (бета).
A-кластер располагается более компактно и занимает участок ДНК размером около 28 тысяч пар нуклеотидов и включает 3 активных гена и 2 псевдогена: ген £ (дзета), псевдоген С (дзета), псевдоген а (альфа) и гены а2 (альфа два) и а, (альфа один).
На разных стадиях эмбриогенеза происходит избирательное и по-следовательное включение и выключение генов А- и В-кластеров. Гены глобина располагаются в А- и В-кластерах в определенной последовательности и транскрибируются слева направо в порядке их экспрессии. У человека образование «взрослого» гемоглобина включает несколько этапов: эмбриональный гемоглобин, гемоглобин плода и гемоглобин взрослого человека (после рождения). Эмбриональный гемоглобин (до 8 недель развития) состоит из двух С, (дзета)- и двух е (эпсилон)-цепей; гемоглобин плода — из двух a-цепей и двух у (гамма)-цепей. После рождения человека гемоглобин состоит из двух а цепей и двух Р цепей, а также 5 (дельта)-цепи (рис. 83).
Гены глобина по внутреннему строению являются мозаичными (см. рис. 83). Мозаичный ген Р-глобина состоит из трех экзонов и двух
интронов, они нумеруются последовательно цифрами (можно буквами). Ген начинается и заканчивается экзоном. Экзоны и интроны транскрибируются и входят в состав первичного РНК-транскрипта (про-мРНК). При процессинге интроны вырезаются и не участвуют в трансляции, мРНК представлена только экзонами, она короче про-мРНК и, соответственно, короче размера своего гена. На концах интронов имеются определенные постоянные последовательности ГТ-АГ (правило ГТ-АГ).
Регуляция экспрессии генов. Прокариоты
У прокариот регуляция экспрессии генов происходит на уровне транскрипции и осуществляется регуляторным геном (ген- регулятор). Ген-регулятор детерминирует синтез регуляторного белка. Регуляторный белок (белок-репрессор) соединяется с оператором, препятствуя соединению РНК-полимеразы с промотором (рис. 84).
Возможны два состояния лактозного оперона: 1 — оперон «выключен», 2 — оперон «включен».
1. Связывание белка-репрессора с оператором прекращает процесс транскрипции: РНК-полимераза не может присоединиться к промотору, экспрессии структурных генов не происходит, ферменты не синтезируются.
2. Поступающий в клетку индуктор (лактоза) соединяется с белком-репрессором, отбирая его от оператора, что обеспечивает присоединение РНК-полимеразы к промотору и экспрессию генов. Результатом этого является синтез белков-ферментов, которые расщепляют лактозу до глюкозы и галактозы.
Лактозный оперон, как единица транскрипции, регулируется по типу негативной регуляции: гены экспрессируются при условии, что они не выключены регуляторным белком (белком-репрессором).
Эукариоты
У эукариот все клетки многоклеточного организма содержат одинаковую ДНК, но в то же время они отличаются друг от друга по морфологическим, физиологическим и биохимическим свойствам. В основе этих фенотипических различий лежит экспрессия разных генов и соответственно синтез разных мРНК и белков. Большая часть белков синтезируется всеми клетками. Для этого необходима экспрессия генов «домашнего хозяйства», они постоянно экспресси-рованы.
Некоторые белки синтезируются только в определенных клетках (глобины и др.), синтез таких белков, а соответственно и экспрессия генов регулируется: гены «включаются» и «выключаются».
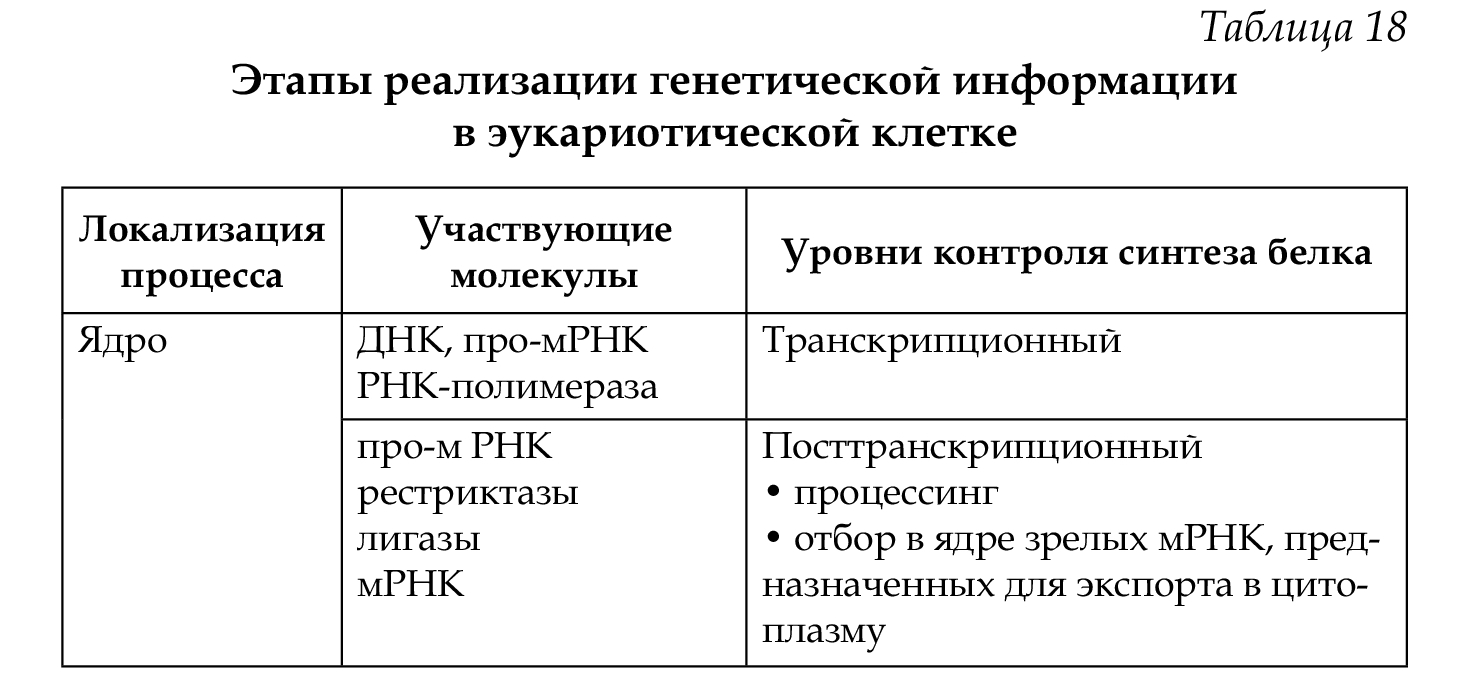
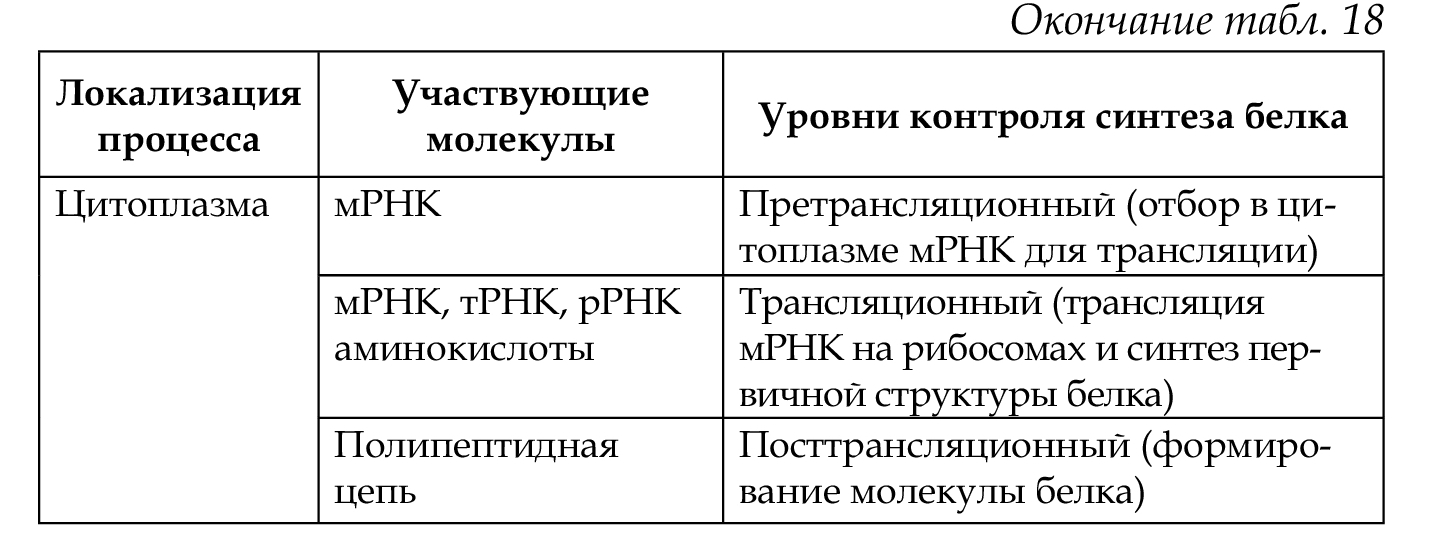
Экспрессия генов регулируется на различных уровнях реализации наследственной информации: претранскрипционном, транскрипционном, посттранскрипционном, претрансляционном, трансляционном и посттрансляционном (табл. 18).
Экспрессия мозаичного гена
Экспрессия мозаичного гена имеет определенные особенности (рис. 85):
• Сложная инициация транскрипции. РНК-полимераза «узнает» промотор и присоединяется к нему после того, как образуется стабильный транскрипционный комплекс (соединение ТАТА- бокса с ТАТА-фактором TF).
• Регулирование скорости и интенсивности транскрипции. Регуляторные белки (сайт-специфические ДНК-связывающие) взаимодействуют с короткими (8-15) нуклеотидными последовательностями в области энхансера (сайленсера), элемента перед промотором, и изменяют процесс транскрипции. Комбинационный эффект регуляторных белков обеспечивает экспрессию гена (+++- -, +- -).
• Сплайсинг. Образование мРНК. Экзоны сплайсируются с образованием мРНК, которая намного меньше, чем сам мозаичный ген. Сплайсинг катализируется сплайсосомами: они узнают определенные последовательности интронов и удаляют их с образованием лассоподобных структур.
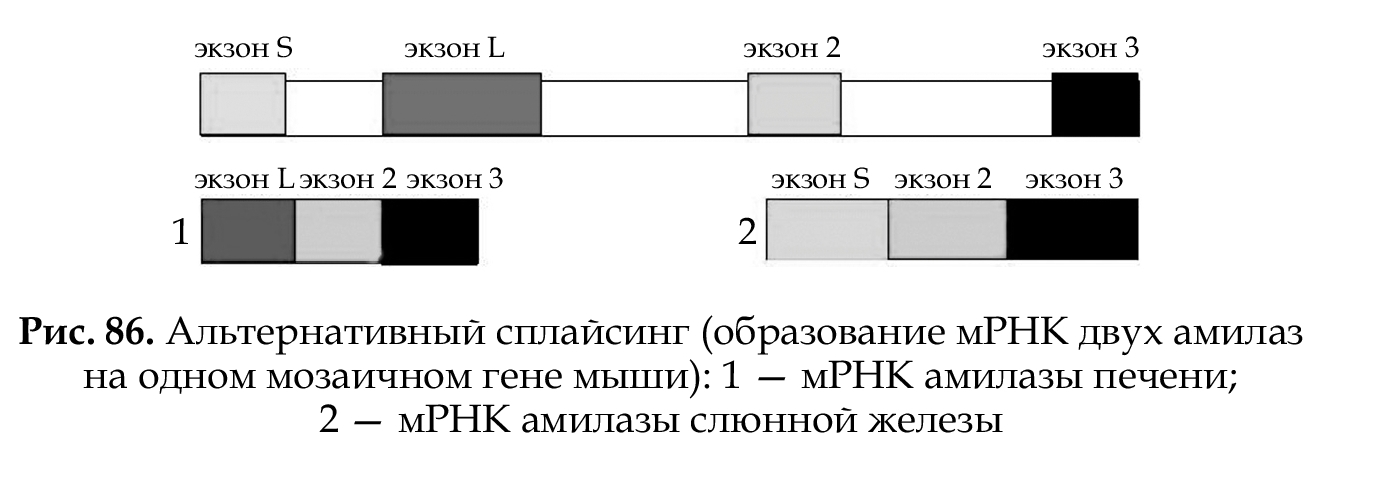
• Альтернативный сплайсинг. Особенности строения мозаичного гена позволяют осуществлять альтернативный сплайсинг: используя одну и ту же генетическую информацию, синтезировать различные изоформы одного белка путем образования различных сочетаний экзонов при их соединении в мРНК (рис. 86).
Экспрессия генов на уровне транскрипции
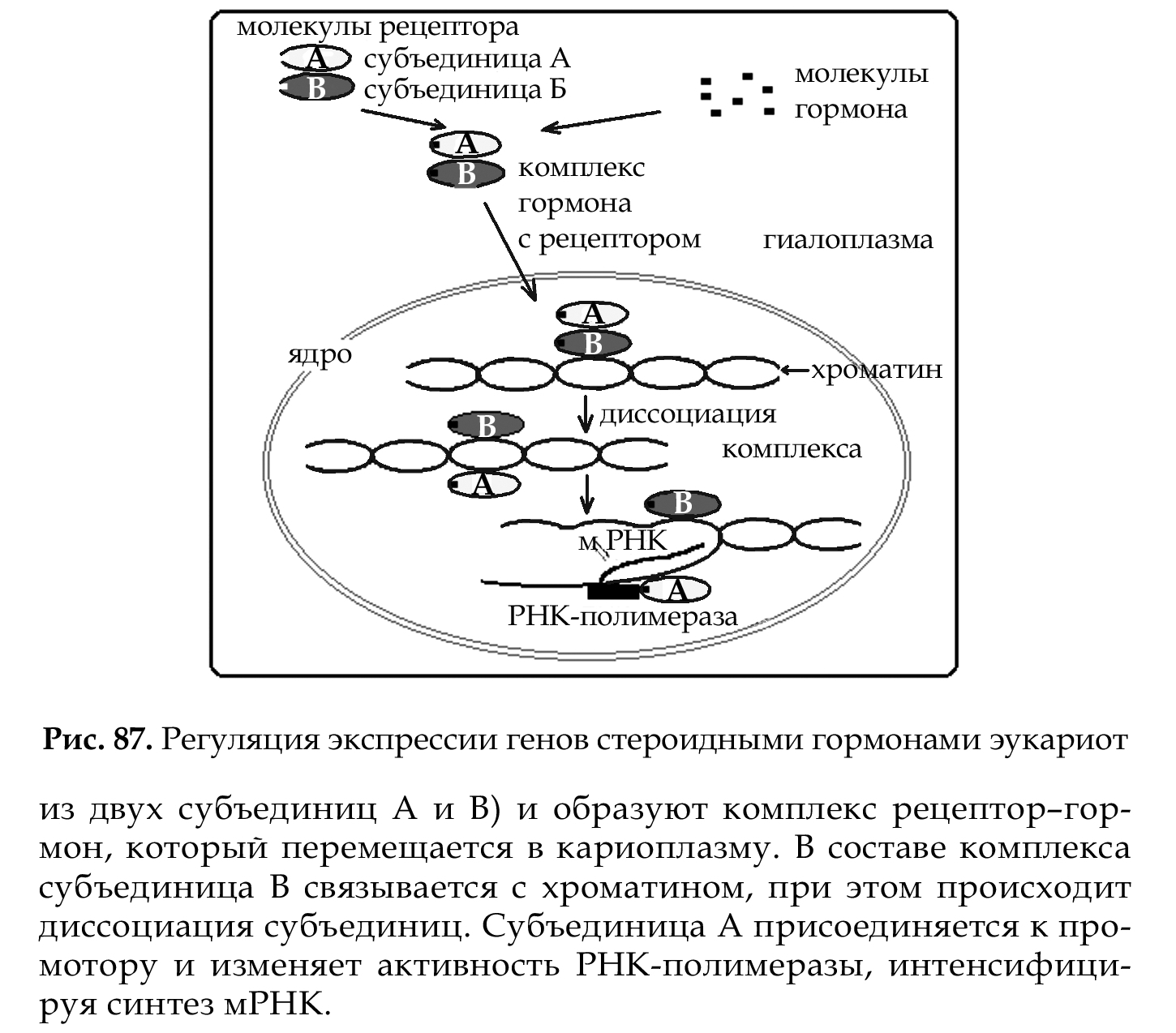
Наиболее изучена регуляция экспрессии генов на уровне транскрипции (рис. 87). Она осуществляется с помощью разнообразных регуляторных белков, а в некоторых клетках и гормонов.
Примером гормональной регуляции экспрессии генов на уровне транскрипции является действие стероидного гормона прогестерона на клетки-мишени. Молекулы гормона поступают в цитоплазму, соединяются с молекулами белка-рецептора (он состоит из двух субъединиц А и В) и образуют комплекс рецептор-гормон, который перемещается в кариоплазму. В составе комплекса субъединица В связывается с хроматином, при этом происходит диссоциация субъединиц. Субъединица А присоединяется к промотору и изменяет активность РНК-полимеразы, интенсифицируя синтез мРНК.