I. Белок - субстрат жизни
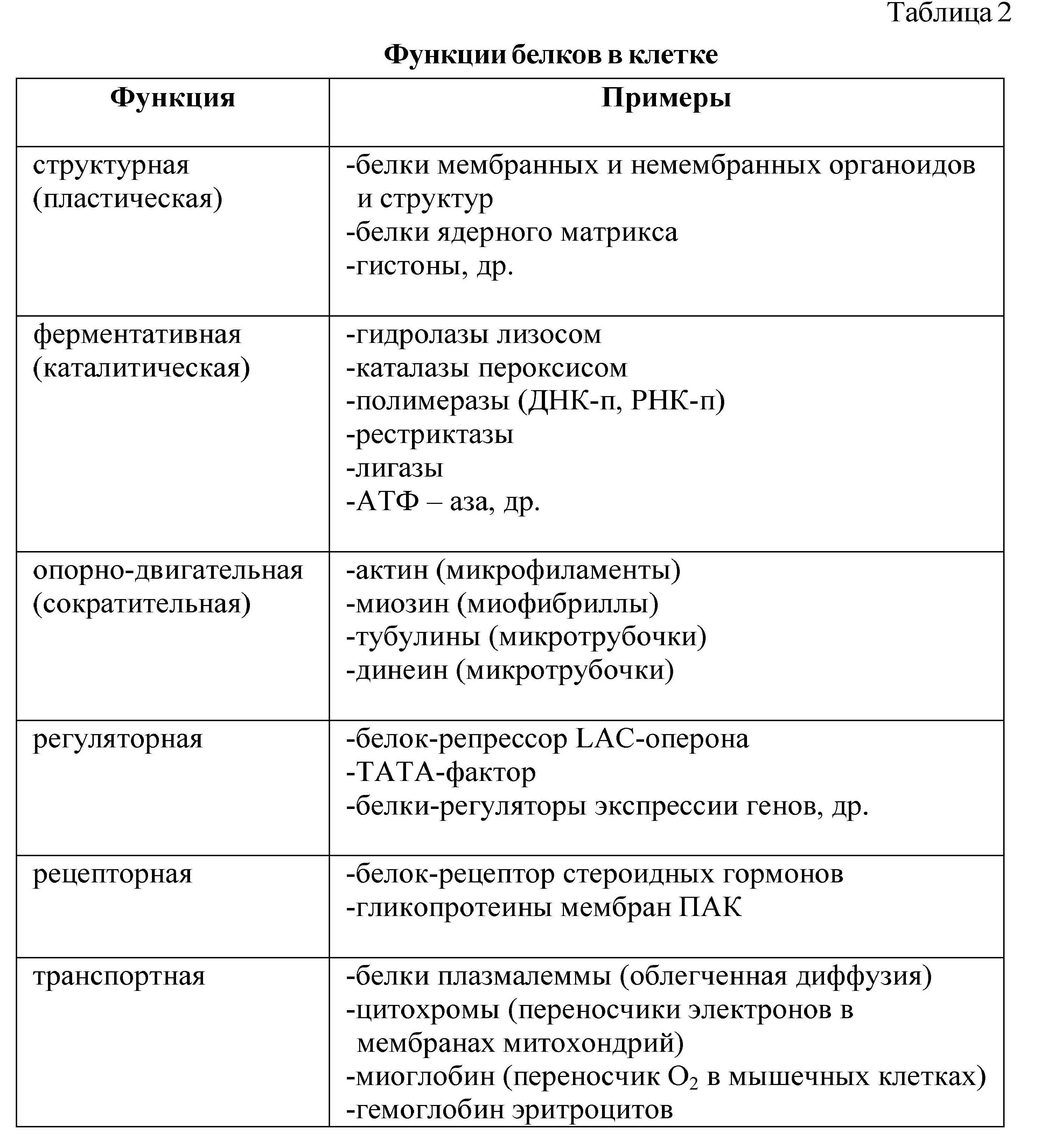
Основу жизнедеятельности живых организмов составляют процессы превращения веществ (окисление, восстановление, расщепление, синтез). В течение жизни каждая клетка усваивает и продуцирует различные вещества, строит и обновляет свои структуры, выполняет определенные функции. Основным строительным материалом в клетке является белок (матриксы цитоплазмы, ядра, митохондрий, пластид; мембранные и немембранные органоиды) - это структурные белки. Особую группу структурных белков составляют сократительные белки, которые формируют опорно-двигательные элементы клетки (микротрубочки, микрофиламенты, микрофибриллы) и определяют движение клеток, деление, фагоцитоз и др. К таким белкам относятся: актин, тубулин, миозин и др. Превращение веществ в клетке осуществляется с помощью ферментов, химической основой которых являются белки. Таким образом, структурная (пластическая) и каталитическая (ферментативная) функции являются главными функциями белка в любой клетке, именно белки определяют и строение клетки и процессы ее жизнедеятельности. Кроме этого, белки выполняют многочисленные функции в клетке и организме (табл. 2).
Белки являются универсальными молекулами и имеют принципиально сходное строение у животных, растений, бактерий и вирусов. Каждый белок в своей первичной структуре представляет собой цепочку аминокислот, соединенных пептидными связями (полипептид). Но в то же время, организмы разных видов различаются своими белками; разные ткани одного и того же организма построены из разных белков (соединительная ткань - коллаген; мышечная ткань - актин, миозин, миоглобин; ногти, волосы - кератин и т.д.);
имеются индивидуальные отличия организмов по строению белков - следовательно, белки обладают специфичностью. Специфичность белков обусловлена особенностью первичной структуры. Полипептидные цепи различаются между собой набором аминокислот, последовательностью их расположения и количеством. Разнообразие белков огромно.
II. Нуклеиновые кислоты
В многоклеточном организме клетки дифференцируются и поэтому, клетки одной ткани сходны, а клетки разных тканей различаются по морфологии и функциям. При делении каждая клетка образует себе подобные дочерние клетки (из клеток печени образуются клетки печени; из клеток кожи - клетки кожи). Чтобы синтезировать белки, характерные для данного типа клеток, чтобы воспроизводить себе подобных - необходимо иметь информацию, заключенную в каком-то материальном субстрате, которую: а) можно использовать в процессе жизнедеятельности и б) передавать дочерним клеткам при делении. Это обеспечивает преемственность в строении и функции клеток и организмов в поколениях. Материальным субстратом - носителем генетической информации является ДНК (у некоторых вирусов - РНК). Реализация генетической информации происходит с участием различных РНК (мРНК, тРНК, рРНК).
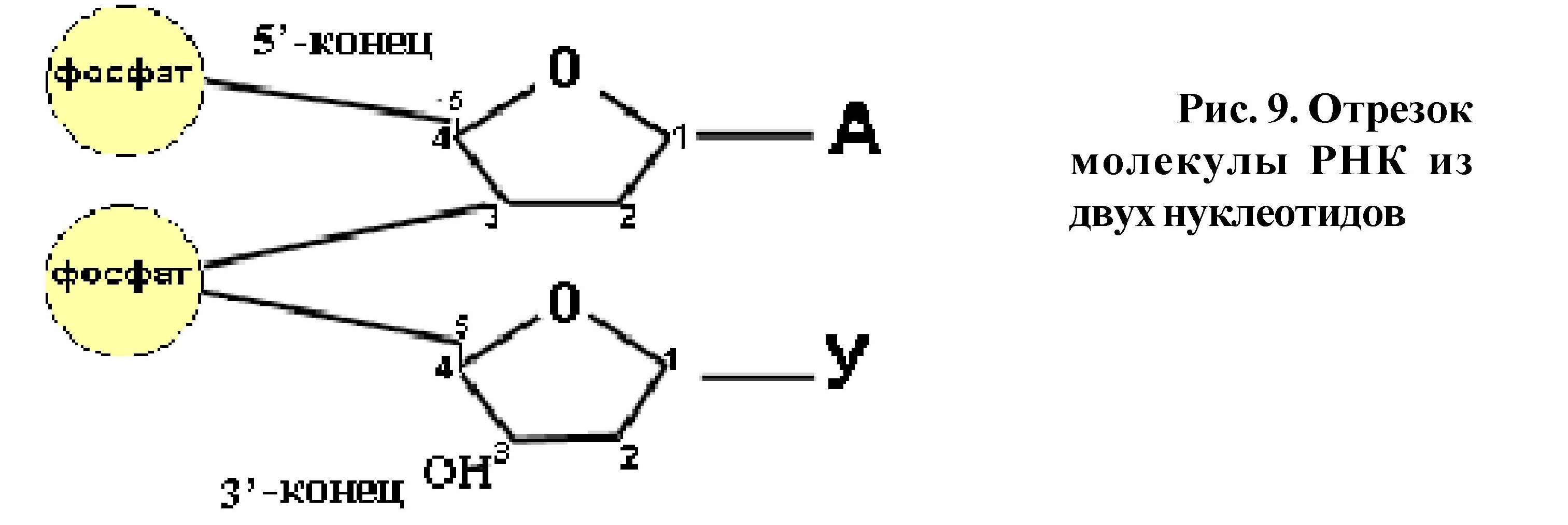
Нуклеиновые кислоты - биополимеры, состоящие из мономеров-нуклеотидов. Любой нуклеотид состоит из трёх частей: углевода, остатка фосфорной кислоты и азотистого основания. Каждая молекула нуклеиновой кислоты - это определённые последовательности нуклеотидов. При соединении нуклеотидов в цепь образуются связи между углеводом и остатком фосфорной кислоты. Углеродный атом в 5 положении рибозы (дезоксирибозы) одного нуклеотида соединяется через фосфатную группу с углеродным атомом в 3 положении сахара предыдущего нуклеотида.
Таким образом, первый нуклеотид в цепи имеет свободный углеродный атом в 5 положении, а последний - в 3 положении (рис. 9), поэтому концы
полинуклеотидных цепей обозначаются как 5/ и 3/. В молекуле ДНК две полинуклеотидные цепи, они антипараллельны, то есть там, где у одной цепи 5/ конец - у второй - 3/ конец и наоборот.
Принципиально строение ДНК и РНК сходно, но есть и отличия: молекула ДНК состоит из двух полинуклеотидных цепей (РНК - одна цепь), в состав ДНК входит углевод дезоксирибоза (РНК - рибоза), в состав ДНК входят азотистые основания - аденин, тимин, гуанин, цитозин (в РНК вместо тимина входит урацил). Нуклеотиды одной цепи ДНК (РНК) различаются между собой только азотистым основанием. Две цепи нуклеотидов ДНК соединяются водородными связями по правилу комплементарности: А-Т; Г-Ц.
ДНК содержится в ядре клетки, РНК - в ядре (в основном, в ядрышке) и цитоплазме (гиалоплазма, рибосомы). Кроме того, некоторые органоиды имеют собственную ДНК и все виды РНК (митохондрии, пластиды).
ДНК была открыта в 1869 году (Мишер), но только в 1953 году было расшифровано строение этой молекулы (Уотсон и Крик).
Основная биологическая роль ДНК состоит в хранении, передаче и самовоспроизведении наследственной (генетической) информации. Строение ДНК универсально (принципиально одинаково у всех живых организмов), но разные молекулы ДНК различаются между собой. Специфичность ДНК зависит от нуклеотидного состава, последовательности нуклеотидов, количества нуклеотидов. Таким образом, от того какие нуклеотиды входят в состав молекулы, как они расположены и сколько их, зависит объём информации и её смысл.
В строении ДНК содержится информация о структуре белков организма и рибонуклеиновых кислот (тРНК, рРНК).
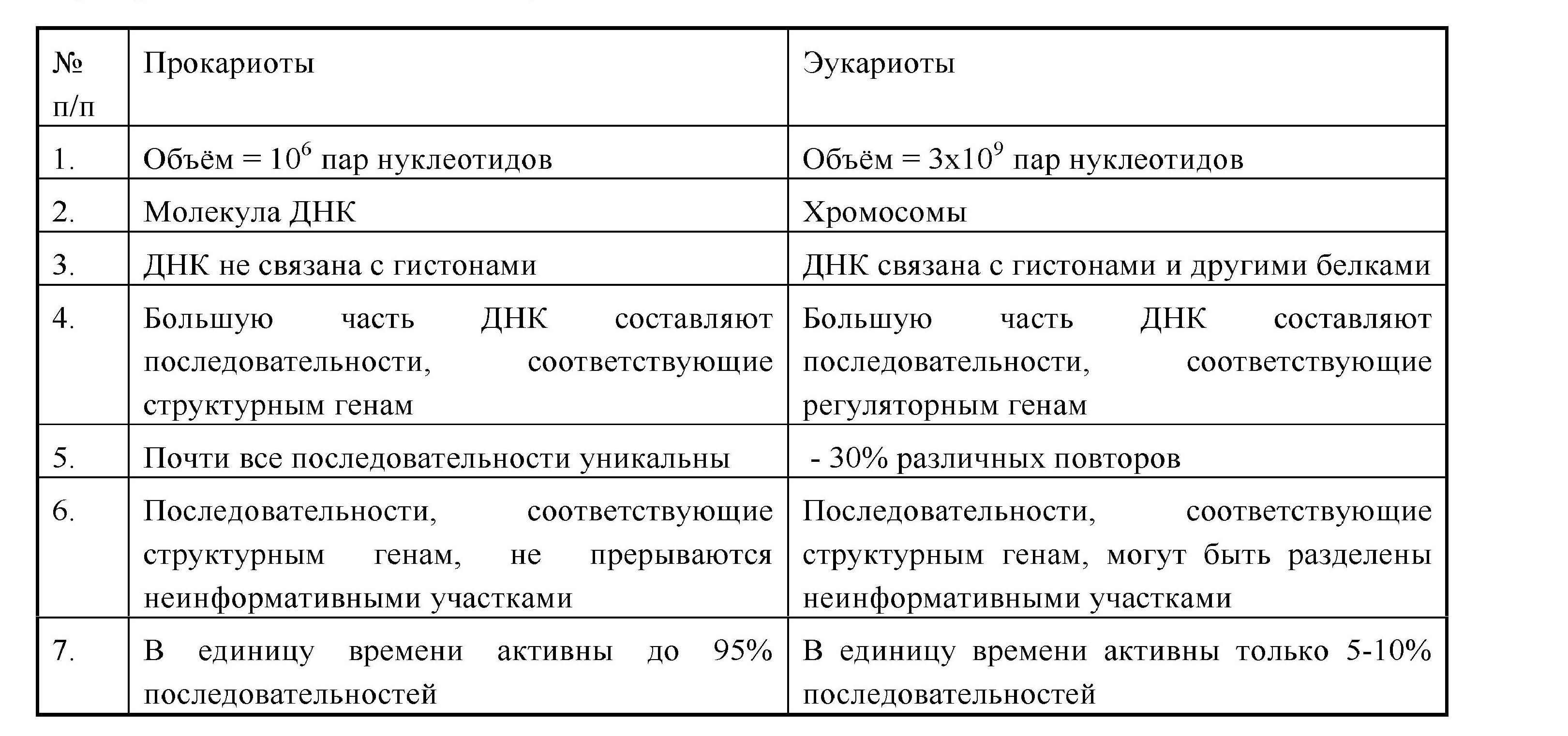
Наследственный аппарат организован по-разному у вирусов, прокариот и эукариот. У вирусов - это может быть молекула ДНК или РНК (различной структурной организации).
У прокариот генетический аппарат представлен двухцепочечной кольцевой молекулой ДНК (нуклеоид, генофор), в которой содержится основная видовая наследственная информация, и плазмоном - совокупностью автономных генетических элементов. Это мелкие кольцевые молекулы ДНК - плазмиды и эписомы, содержащие ограниченную информацию о некоторых признаках данного организма (в плазмидах R находятся гены устойчивости к антибиотикам;эписомы F определяют способность к размножению). Плазмиды и эписомы способны к репликации и перемещению из клетки в клетку при конъюгации.
У эукариот генетический аппарат представлен надмолекулярными структурами - хромосомами, химической основой которых является хроматин (ДНК + белки). Хроматин может быть конденсирован, неактивный - гетерохроматин, или деконденсирован, активный - эухроматин (см. стр. 24). Не вся ДНК эукариот является информативной. Большая часть ее представлена
регуляторными последовательностями. Многие участки повторяются в геноме (умеренные и высокие повторы).
Основные различия в организации генетического материала у про- и эукариот сведены в таблицу 3.
III. Генетический код, его характеристика
Смысл генетической информации зашифрован в молекуле ДНК. Генетический код - это система записи генетической информации, которая используется клеткой в процессе жизнедеятельности. Другими словами - это система расположения нуклеотидов в молекуле ДНК, определяющая последовательность аминокислот в молекуле белка (правило коллинеарности). Единицей генетического кода является триплет нуклеотидов в молекуле ДНК (кодон), который соответств ует одной аминокислоте.
Генетический код характеризуется:
а) универсальностью (другого способа записи генетической информации в природе нет)
б) триплетностью (единица генетического кода - триплет нуклеотидов - кодон)
в) избыточностью (вырожденностью)
г) однозначностью
д) наличием смысловых, терминирующих и инициирующих кодонов.
IV. Реализация генетической информации в клетке
Реализация генетической информации происходит в течение всей жизни клетки в процессе биосинтеза белков, характерных для данного вида организмов (клеток).
Интенсивность биосинтеза белка наибольшая в интерфазе, снижается к началу деления, почти нулевая при делении и возрастает сразу после деления. Биосинтез белка можно разделить на два этапа: транскрипция (происходит в ядре на ДНК) и трансляция (происходит в цитоплазме на рибосомах).
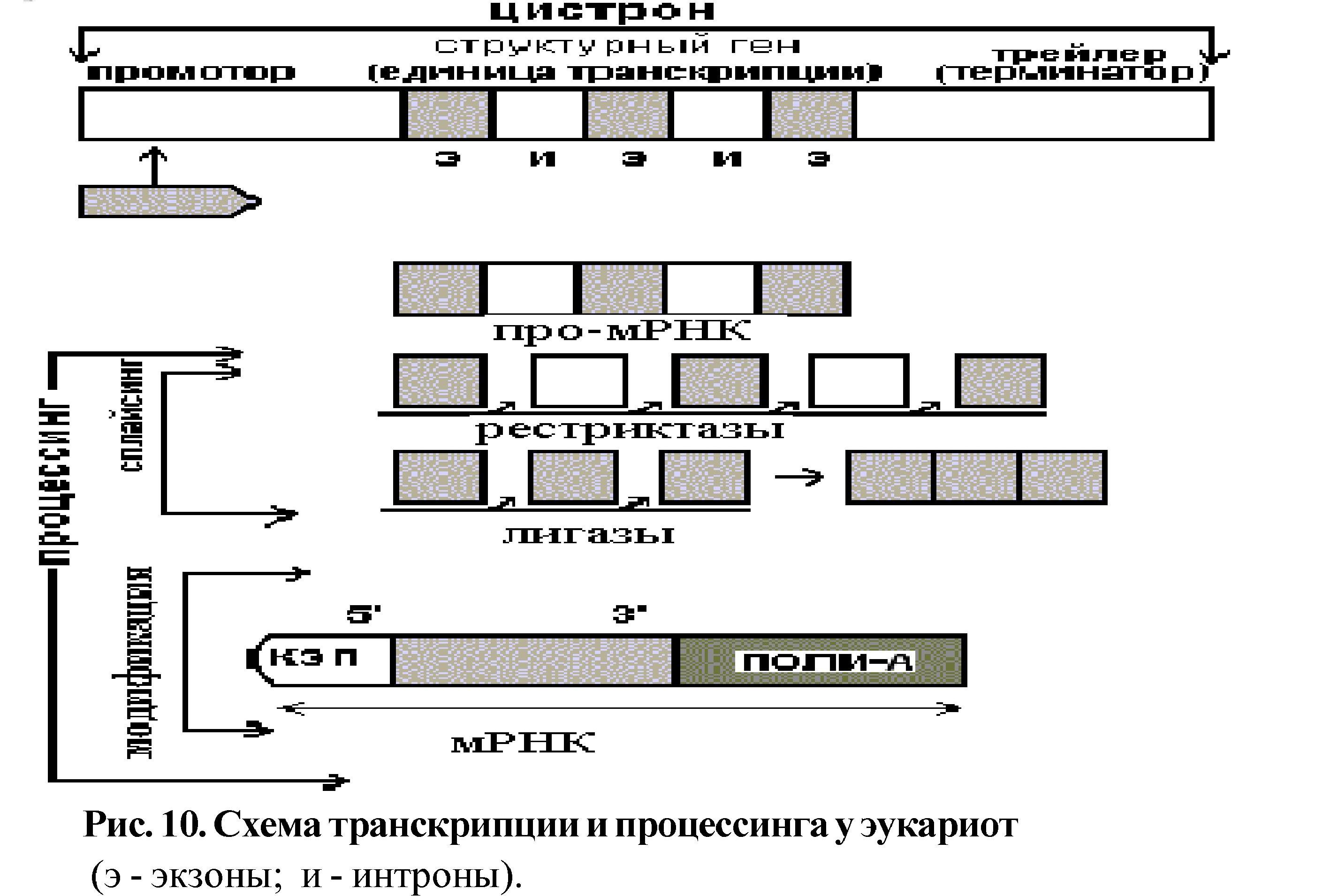
Функциональной единицей, которая участвует в транскрипции, является цистрон - отрезок ДНК состоящий из трёх частей:
а) промотор (около 40 пар последовательностей), с которым связывается фермент РНК-полимераза;
б) последовательности, соответствующие структурному гену;
в) терминальный участок (трейлер), где заканчивается транскрипция.
Биологической сущностью транскрипции является "переписывание" генетической информации с молекулы ДНК на РНК, а химической - синтез молекулы мРНК. Биологической сущностью трансляции является перевод информации с языка нуклеотидов на язык аминокислот (расшифровка генетического кода), а химической - синтез полипептидной цепочки.
Оба процесса относятся к реакциям матричного синтеза, которые характеризуются: наличием молекулы-образца (матрицы), энергии, специфического фермента, выполнением правила комплементарности и протекают в три этапа (инициация, элонгация, терминация).
У прокариот синтезированная мРНК сразу может служить матрицей для трансляции.
У эукариот информативные последовательности структурного гена (экзоны) разделены неинформативными (интроны). Первичный транскрипт включает как экзоны, так и интроны - это незрелая или про-мРНК. Затем начинается процессинг (созревание мРНК), в результате которого удаляются интроны и образуется зрелая мРНК, состоящая только из экзонов. Процессинг состоит из двух этапов: сплайсинга и модификации (рис. 10).
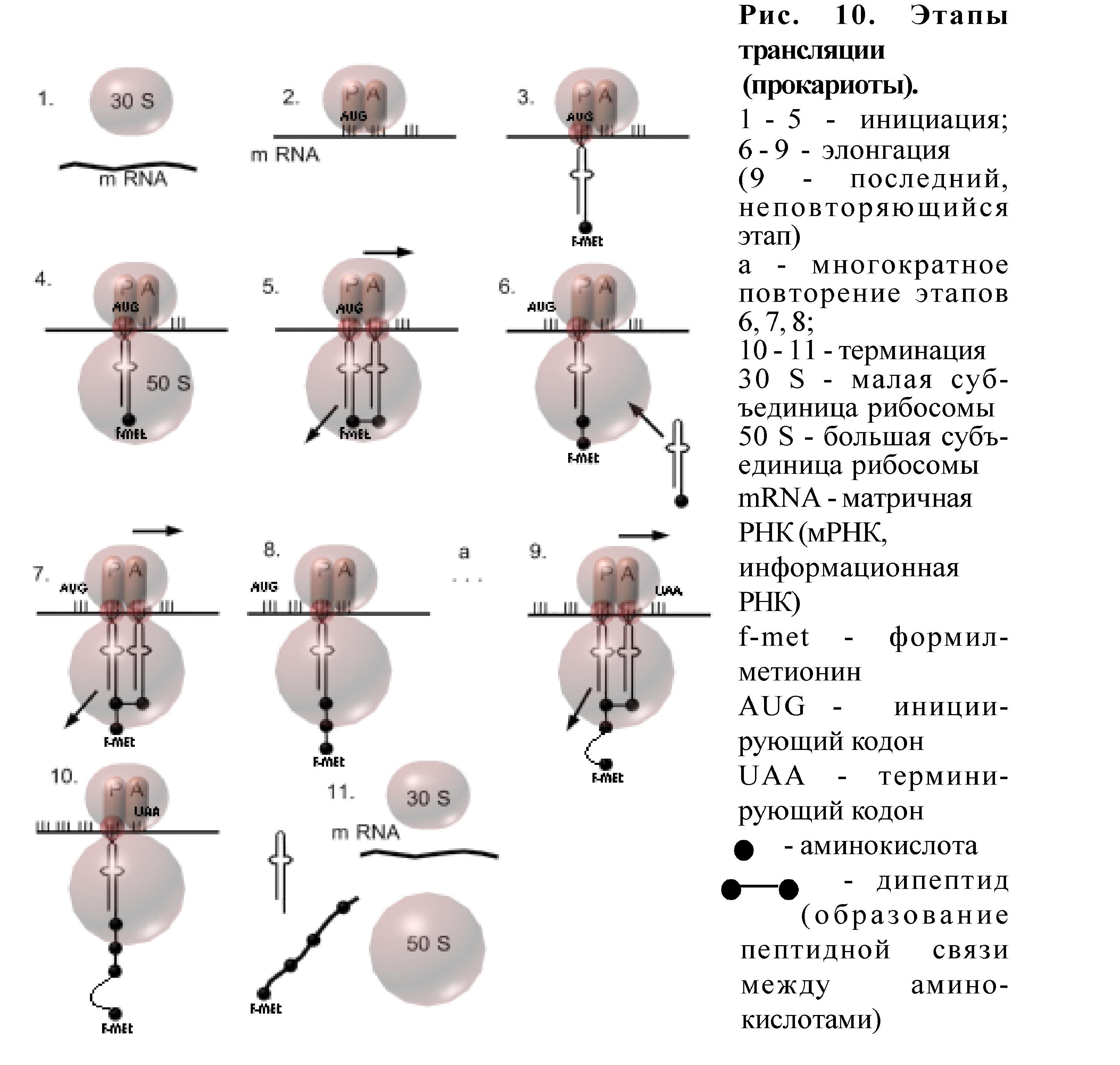
Трансляция (рис.10) также состоит из инициации, элонгации и терминации.
Местом трансляции являются рибосомы. В рибосомах есть два активных функциональных центра - пептидильный и аминоацильный. Не работающая рибосома диссоциирована на две субъединицы: малую и большую.
Инициация трансляции начинается со связывания мРНК с малой субъединицей рибосомы, причём необходимо чтобы в её пептидильном центре оказался триплет АУГ - это инициирующий кодон. С этим кодоном связывается тРНК-f-метионин, а затем малая и большая субъединицы рибосомы объединяются. Рибосома готова к работе. В аминоацильном центре рибосомы находится другой триплет нуклеотидов мРНК, с которым может связаться тРНК, имеющая комплементарный антикодон. Когда это произойдёт, то между двумя аминокислотами (одна - f-метионин в пептидильном центре, вторая - в аминоацильном центре) возникает пептидная связь - образуется дипептид, инициация завершилась. Рибосома передвигается по мРНК на один триплет, который оказывается в аминоацильном центре, тРНК из него перемещается в пептидильный центр; она связана с дипептидом, а первая тРНК уходит в цитоплазму. Аминоацильный центр свободен, в нем находится новый кодон, с которым может связаться тРНК с комплементарным антикодоном. Так, передвигаясь по мРНК, рибосома "прочитывает" информацию, переводит её на язык аминокислот и полипептидная цепь наращивается. Это - элонгация. Элонгация происходит до тех пор, пока на пути рибосомы в А-центре не окажется кодон-терминатор. Тогда полипептидная цепь отсоединяется от рибосомы, мРНК тоже отделяется от рибосомы, рибосома диссоциируется на субъединицы, происходит терминация
Этапы трансляции (рис.10):
Инициация 1-5. Начало матричного синтеза (трансляции) происходит поэтапно:
1- связывание мРНК с малой (30 S) субъединицей рибосомы
2 - установка в пептидильном центре (Р) инициирующего кодона АУГ (AUG)
3 - связывание тРНК с аминокислотой формил-метионин (тРНК - f-met) с кодоном АУГ (образование инициирующего комплекса)
4 - присоединение большой (50 S) субъединицы рибосомы
5.1-образование комплекса кодон-антикодон в аминоацильном
(А) центре
5.2 - образование пептидной связи между формил-метионином и второй аминокислотой(образование дипептида)
5.3 - транспозиция рибосомы (перемещение) по мРНК на один триплет (при этом первая тРНК покидает рибосому, вторая тРНК, с которой связан дипептид, перемещается из А - в Р центр, а в А центре появляется новый кодон).
Элонгация 6 - 9. Углубление и ускорение процесса трансляции, результатом чего является наращивание полипептидной цепи. Состоит из многократно повторяющихся этапов:
6 - транспортировка аминокислот в рибосому с помощью тРНК
7.1-образование комплекса кодон-антикодон в А центре
7.2 - образование пептидной связи между аминокислотами
8 - транспозиция рибосомы по мРНК на один триплет
Терминация 10-11. Окончание трансляции.
10 - появление в А центре после очередной транспозиции рибосомы терминирующего кодона (УАА, УАГ, УГА)
11.1-в Р- центре дестабилизируется и утрачивается связь между тРНК и мРНК
11.2 - полипептид отщепляется от тРНК
11.3 - мРНК покидает рибосому
11.4 - диссоциация рибосомы на субъединицы
V. Репликация ДНК
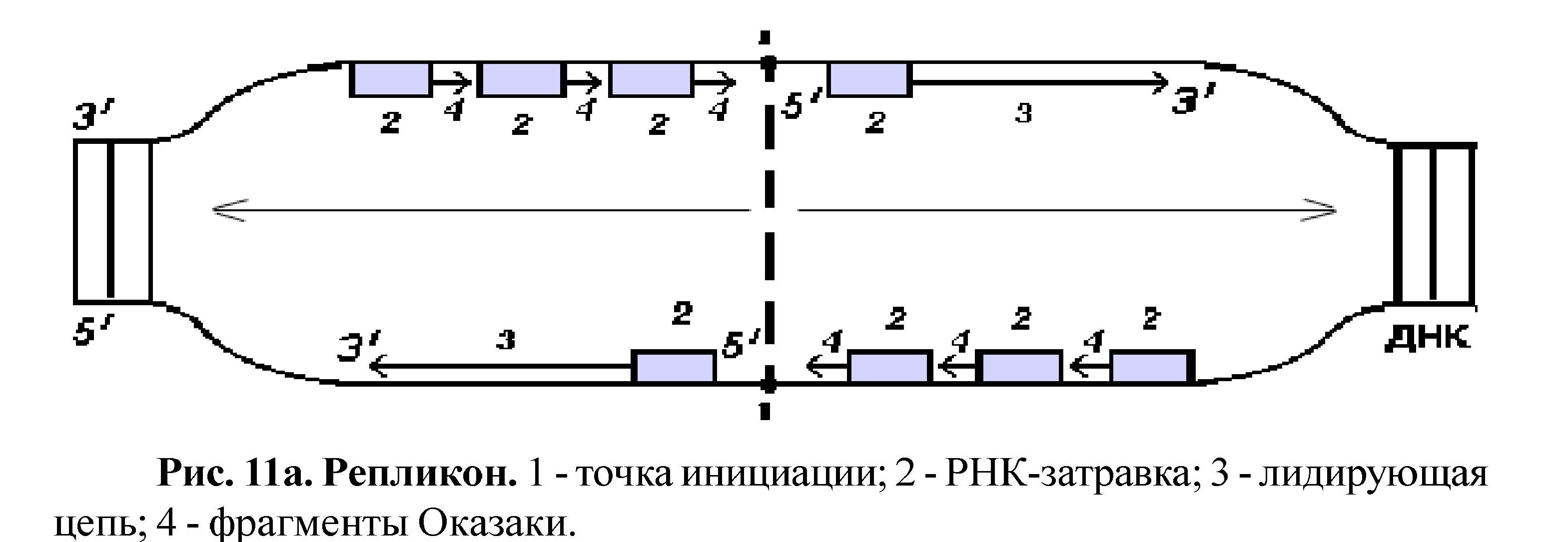
Самовоспроизведение (ауторепродукция) ДНК называется репликацией. Репликация ДНК происходит перед делением клетки; в результате этого процесса содержание ДНК в клетке удваивается, а так как репликация протекает по правилу комплементарности, то две дочерние молекулы идентичны материнской и друг другу. Следовательно, каждая новая клетка получает информацию в количественном и качественном отношении одинаковую с родительской клеткой. Разъединение двух цепей ДНК у эукариот начинается одновременно в нескольких участках (у прокариот в одном месте). Такой участок называется - репликон (рис. 11а). В эукариотической клетке может быть более 2000 репликонов. Репликация - это реакция матричного синтеза; матрицей служит молекула ДНК, основными ферментами являются ДНК-полимераза, лигаза, рестриктаза.
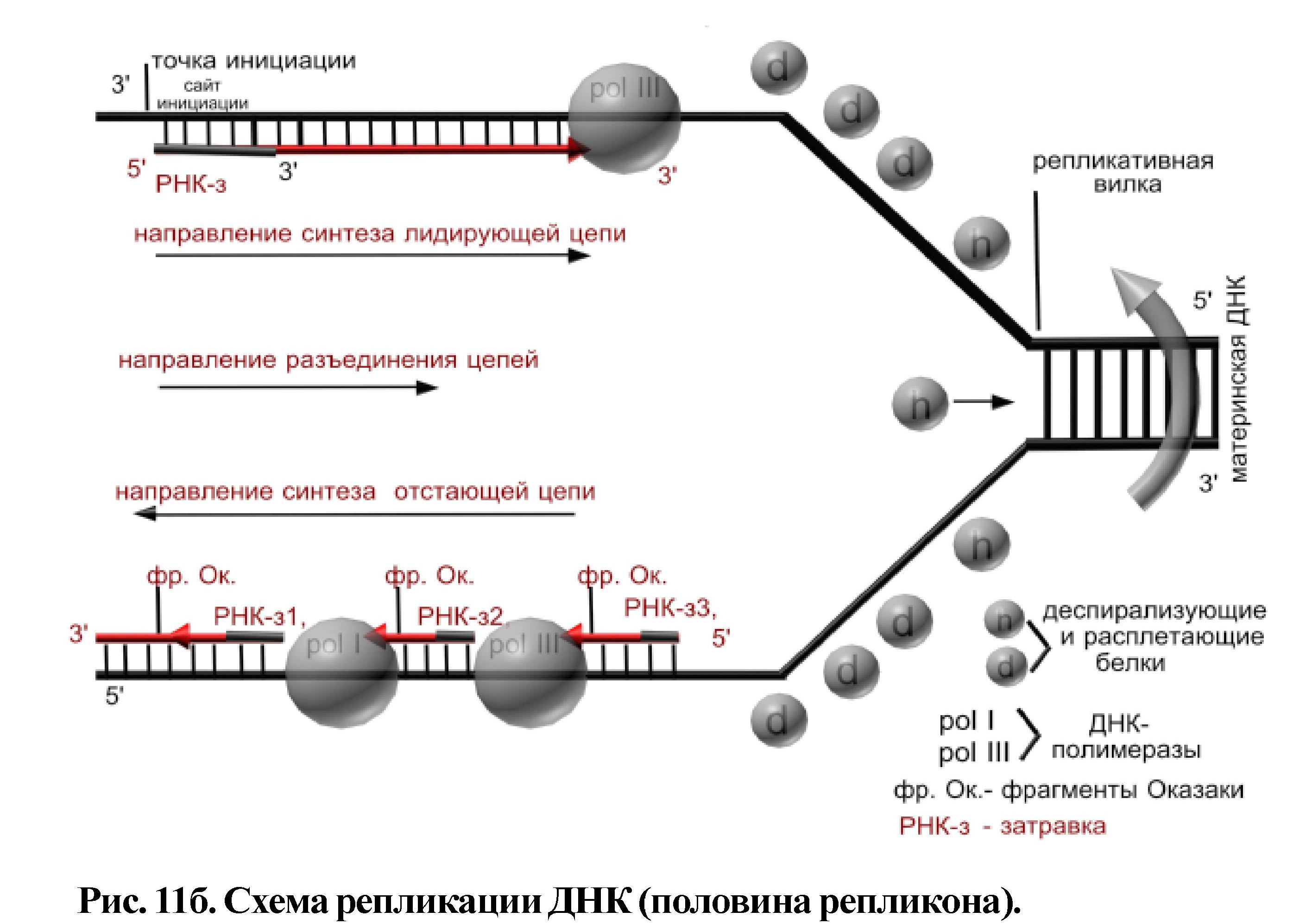
Начинается процесс с разрыва водородных связей между азотистыми основаниями ДНК на участке, включающем около 300 пар нуклеотидов - это место называется точка инициации. Так как разъединение цепей ДНК от точки инициации идет вправо и влево одновременно, цепи ДНК антипараллельны, а фермент ДНК-полимераза может работать только в одном направлении (соединяя нуклеотиды от 5 углерода последующего к 3 углероду предыдущего), то синтез дочерних цепей идет по-разному на разных участках одного репликона. Одна цепь - лидирующая, синтезируется непрерывно, а вторая - отстающая, синтезируется фрагментарно (Рис. 11б).
На цепи 3/_5/ рядом с точкой инициации есть особая последовательность нуклеотидов - сайт инициации, на котором синтезируется небольшая молекула РНК (РНК-затравка). У РНК-затравки свободен 3/ - конец, к которому присоединяется первый нуклеотид ДНК, к нему второй и т. д. В результате синтезируется лидирующая дочерняя цепь. На противоположной, антипараллельной цепи (5/-3/) сайта инициации нет и проходит время, пока в
результате разрыва водородных связей обнаружится такой сайт; РНК-затравка синтезируется и от неё в сторону противоположную направлению разъединения ДНК синтезируется небольшой фрагмент дочерней цепи. После разъединения следующего участка молекулы ДНК, следующая молекула РНК-затравка находит свой сайт и синтезируется новый фрагмент дочерней цепи ДНК в направлении 5/ -3/ и т.д. Таким образом, эта цепь синтезируется небольшими фрагментами (фрагменты Оказаки) и отстаёт во времени. На другой половине репликона, где разъединение цепей ДНК идёт в другую сторону, также, в одном направлении дочерняя цепь синтезируется непрерывно, в другом - фрагментарно. Затем рестриктазы вырезают РНК-затравки (одну - из лидирующей цепи и от каждого фрагмента Оказаки на отстающей цепи), ДНК-полимераза достраивает молекулу ДНК на местах вырезанных РНК-затравок, а лигазы соединяют фрагменты в непрерывную цепь. В каждой новой молекуле ДНК одна цепь старая (материнская), а вторая - новая (дочерняя). Такой способ репликации называется полуконсервативным.
VI. Обратная транскрипция
Представление о направлении потока информации в клетке и последо-вательности процессов получило название центральной догмы молекулярной биологии. Передача генетической информации идёт в направлении
Однако, оказалось, что иногда информация может передаваться от РНК к ДНК. Это явление было изучено у вирусов, генетический аппарат которых представлен не ДНК, а РНК. Это группа ретровирусов, к которым относится вирус гриппа, СПИДа и др. Чтобы после внедрения таких вирусов в клетку хозяина их генетическая информация могла быть использована для синтеза вирусных белков, необходимо на вирусной РНК синтезировать ДНК, с последующим встраиванием ее в геном клетки. Этот процесс идёт под контролем фермента ревертазы (обратной транскриптазы) и называется обратной транскрипцией. Таким образом, направление потока генетической информации в клетке в окончательном виде выглядит так:
Открытие явления обратной транскрипции сыграло большую роль в развитии генной инженерии, микробиологии. С помощью ревертаз получают важные лекарственные препараты белковой природы (интерферон, гамма- глобулин и др.), вводя в микробную клетку мРНК человека с информацией о строении этих белков.