I. Ген, классификация и организация генов
Генетика изучает универсальные для всех живых организмов закономерности наследственности и изменчивости.
Элементарными дискретными единицами наследственности являются гены. Воспроизведение и действие генов непосредственно связано с матричными процессами. В настоящее время ген рассматривается как единица функционирования наследственного материала. Химической основой гена является молекула ДНК.
Существует несколько подходов к классификации генов, каждый из которых отражает особенности их функционирования в процессе онтогенеза. Гены, как единицы функции наследственного материала, разделяются на структурные, регуляторные и гены-модуляторы.
Структурные гены содержат информацию о структуре белка (полипептидов) и рибонуклеиновых кислот (рибосомальной и транспортной), при этом генетическая информация реализуется в процессе транскрипции и трансляции или только транскрипции.У человека насчитывается около 30 000 структурных генов, но только часть из них экспрессирована.
Жизнедеятельность клеток обеспечивается небольшим набором функционирующих генов, среди них различают гены «домашнего хозяйства» - ГОФ (гены общеклеточных функций)и гены «роскоши» - ГСП (гены специализированных функций). ГОФ обеспечивают осуществление универсальных клеточных функций, которые необходимы для деятельности всех клеток (гены гистонов, гены р-РНК и т-РНК и др.). ГСП: 1- избирательно экспрессируются в специализированных клетках, определяя их фенотип (гены глобинов, иммуноглобулинов и др.); 2 - функционируют при определенных условиях внешней среды и представляют гены “адаптивного ответа”. Принадлежность к ГОФ или ГСП определяется структурой инициатора.
Регуляторные гены (ген - регулятор лактозного оперона, ген TFM и др.) координируют активность структурных генов на уровне клетки, а также дерепрессию и репрессию генов на уровне организма. Наряду с регуляторными генами, имеются регуляторные последовательности (промотор, оператор, терминатор, энхансеры, сайленсеры, элемент перед промотором), функция которых выявляется во взаимодействии со специфическими белками.
Гены-модуляторы усиливают или ослабляют действие структурных генов, изменяя их функциональную активность.
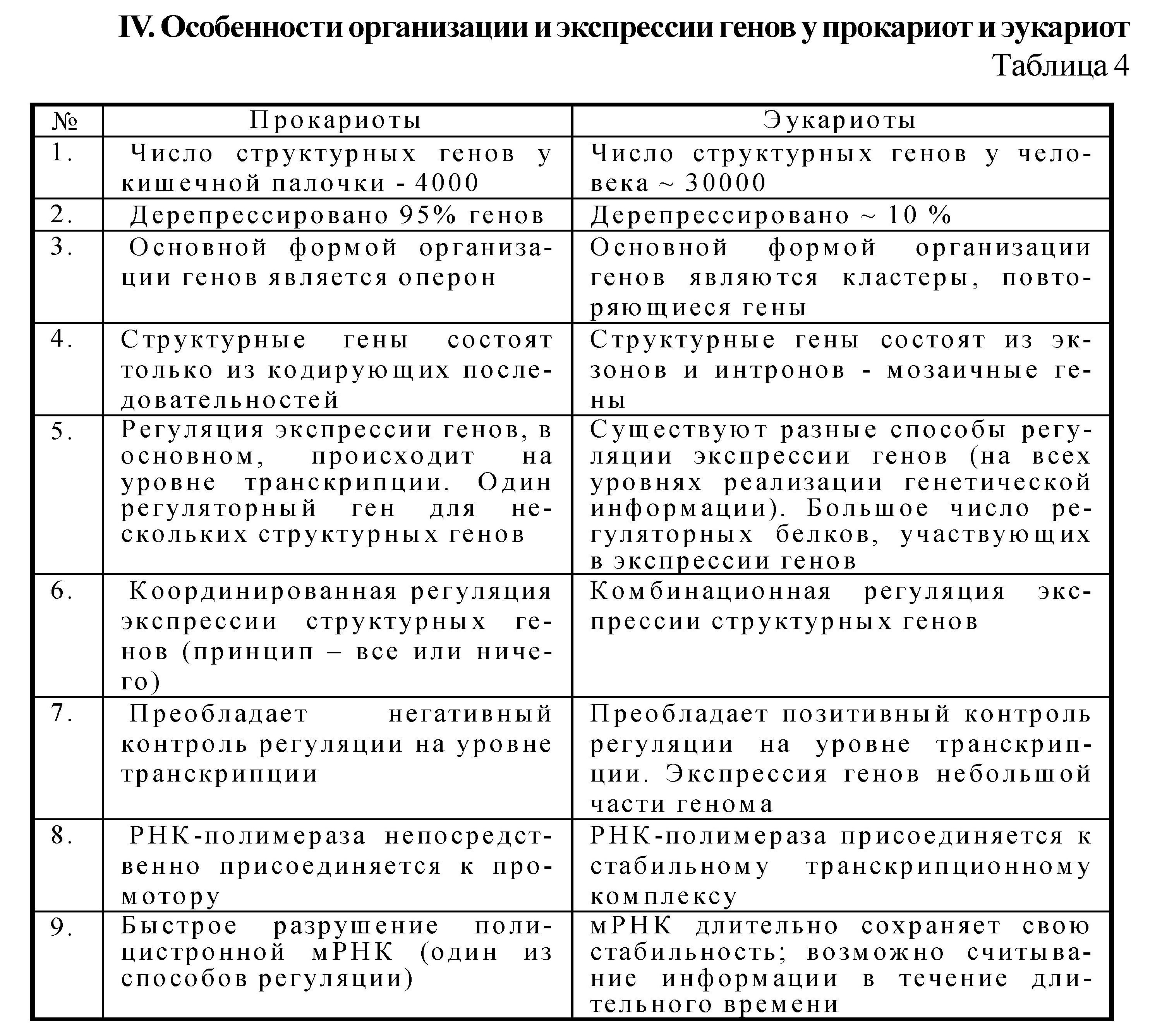
Структурные гены различным образом организованы у прокариот и эукариот.
У прокариот структурные гены организованы в виде независимых генов, транскрипционных единиц и оперонов.
Независимые гены состоят из непрерывной последовательности кодонов, они постоянно экспрессированы и не регулируются на уровне транскрипции (ген-регулятор лактозного оперона). Транскрипционные единицы - группы разных генов, которые связаны функционально и транскрибируются одновременно, что обеспечивает в последующем одинаковое количество синтезируемых продуктов. Обычно это гены белков или нуклеиновых кислот (у кишечной палочки в составе одного из транскриптонов находятся два гена т-РНК, три гена р-РНК).
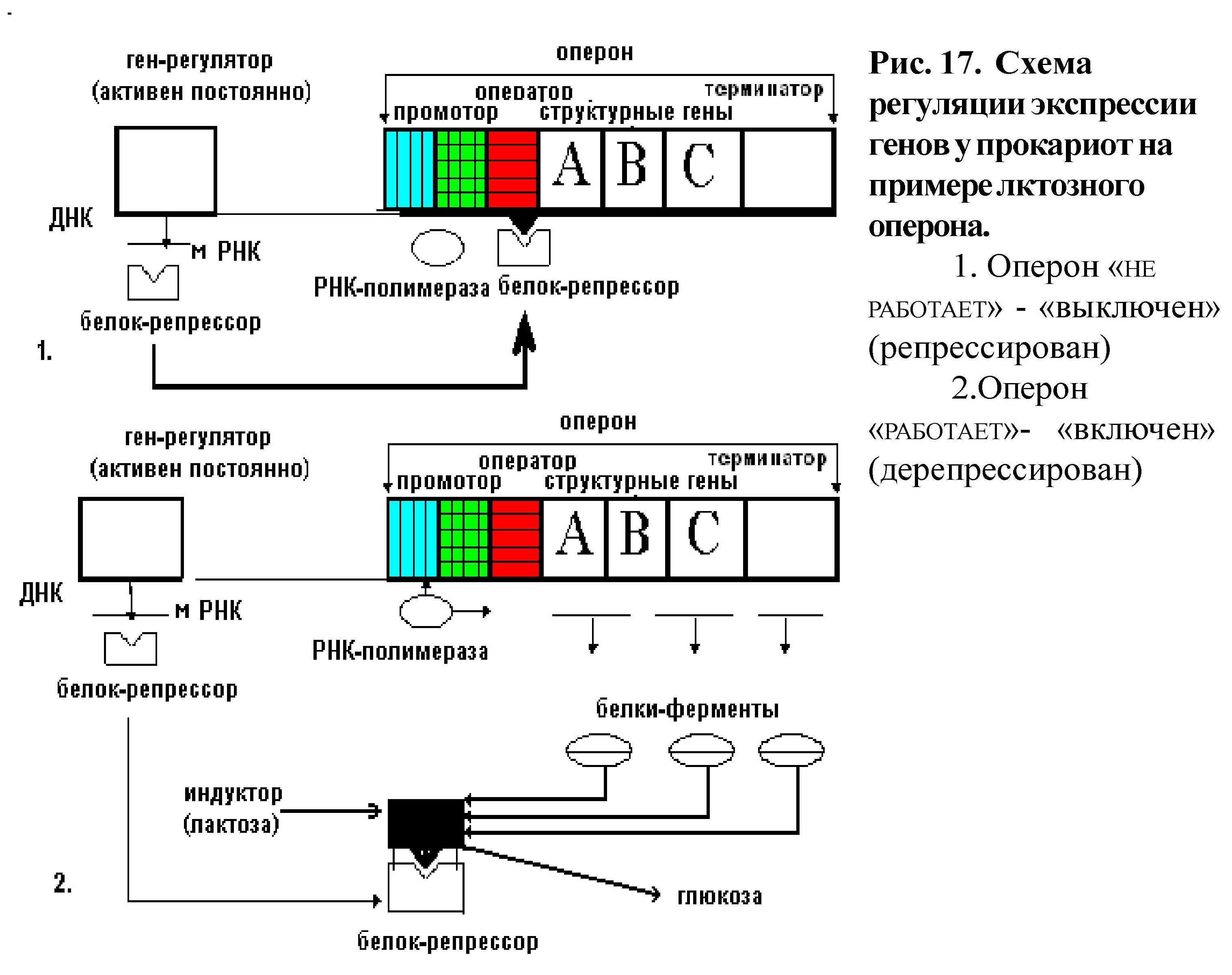
Оперон - это группа структурных генов, следующих друг за другом, находящихся под контролем оператора - определённого участка ДНК. Структурные гены имеют общий промотор, оператор и терминатор, участвуют в одном метаболическом цикле и регулируются координированно (рис. 17).
У эукариот структурные гены, функция которых связана с регуляторными, организованы в виде независимых генов, повторяющихся генов и кластеров генов.
Независимые гены, как правило, располагаются индивидуально, их транскрипция не связана с транскрипцией других генов. Активность некоторых из них регулируется гормонами.
Повторяющиеся гены присутствуют в хромосоме в виде повторов (копий) одного гена - гены гистонов, тРНК, рРНК. Причина повторяемости гистоновых генов определяется необходимостью синтезировать большое количество гистонов, которые являются основными структурными белками ядра (суммарная масса гистонов равна массе ДНК).
Кластер генов - это группа различных генов с родственными функциями, локализованных в определённых участках хромосом. В состав кластера входят активно функционирующие гены и псевдогены ( Нуклеотидные последовательности псевдогенов сходны с последовательностями функционально активных генов, но псевдогены не экспрессируются и не образуют белок. Часто кластеры являются семейством генов, произошедшим от некоего гена-предка.
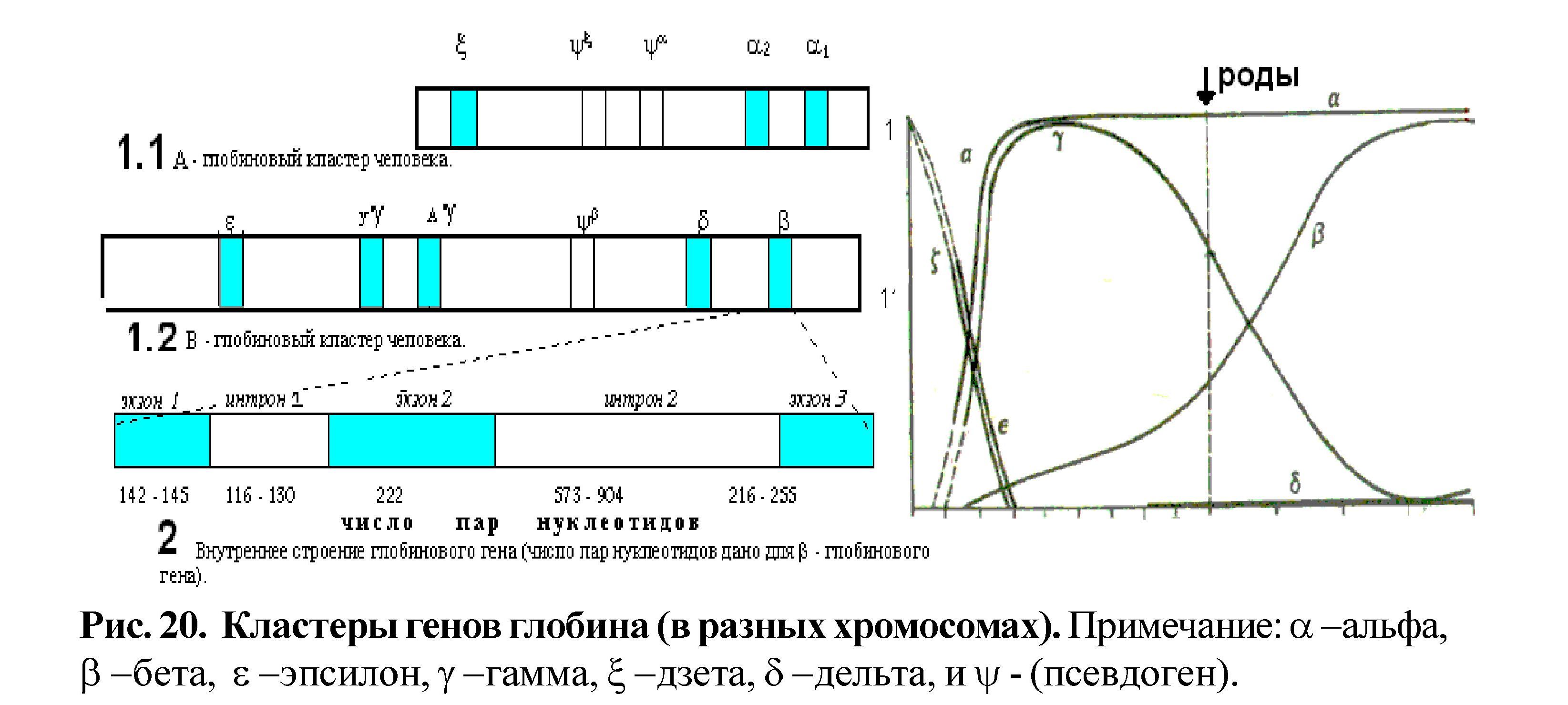
Классическим примером являются гены глобина в составе А и В кластеров. Гемоглобин представлен гемом и белком тетрамером-глобином. Глобиновый тетрамер состоит из двух идентичных цепей и двух идентичных цепей. Аминокислотная последовательность каждой глобиновой цепи и кодируется своим собственным геном, входящим соответственно в состав А или В кластера. У человека А кластер располагается в 16 хромосоме, а В кластер в 11 хромосоме (рис. 20). Кластер В занимает участок ДНК размером 50 тысяч пар нуклеотидов и включает в себя пять функционально активных генов и один псевдоген: ген (эпсилон); два гена (гамма); псевдоген (бэта); ген (дельта) и ген (бэта).
Кластер А располагается более компактно и занимает участок ДНК размером более 28 тысяч пар нуклеотидов и включает в себя активный ген (дзета), псевдоген (дзета), псевдоген (альфа) и гены (альфа) два и (альфа) один, кодирующие идентичные белки. Гены глобина по внутреннему строению являются мозаичными.
Повторяющиеся гены и кластеры глобиновых генов относятся к мультигенным семействам.
II. Регуляция генов у прокариот и эукариот
У про- и эукариот различные варианты организации генов и формы хранения наследственной информации, что предполагает и обеспечивает различный уровень регуляции экспрессии генов в процессе реализации генетической информации.
Для прокариот характерна регуляция на уровне транскрипции. Теория генетической регуляции белкового синтеза была разработана генетиками Ф. Жакобом и Ж. Моно. Объектом изучения служил лактозный оперон кишечной палочки (рис. 17). Кишечная палочка может размножаться в среде, содержащей как глюкозу, так и лактозу. Ферменты для усвоения глюкозы присутствует в клетке постоянно (конститутивные ферменты), а ферменты, необходимые для усвоения лактозы, синтезируется только при её поступлении в клетку (адаптивные ферменты). Синтез адаптивных ферментов регулируется. Лактозный оперон состоит из промотора, оператора (нуклеотидные последовательности которых перекрываются), трёх структурных генов, расположенных друг за другом, и терминатора. Структурные гены детерминируют синтез ферментов, которые необходимы для одного метаболического цикла (расщепление лактозы до глюкозы), при этом каждый ген определяет синтез одного белка-фермента. Оператор регулирует активность структурных генов, включая и выключая их из процесса транскрипции. Сам оператор контролируется геном-регулятором, который располагается на некотором расстоянии от оперона. Ген-регулятор кодирует синтез особого белка-репрессора, который обладает аллостерическим эффектом, что позволяет ему соединяться или с опероном или с поступающей в клетку лактозой. Гены оперона функционируют совместно, поэтому одновременно синтезируются все три фермента или не синтезируется ни одного.
Могут быть два состояния оперона: оперон «выключен» и оперон «включен» и соответственно два положения белка-репрессора.
Оперон «выключен» (рис. 17.1): белок-репрессор соединен с оператором. РНК-полимераза не может присоединиться к промотору, транскрипции структурных генов и синтеза ферментов не происходит.
Оперон «включен» (рис. 17.2): лактоза поступает в клетку и соединяется с белком-репрессором, отбирая его от оператора, при этом оператор освобождается и РНК-полимераза соединяется с промотором, осуществляя транскрипцию структурных генов. Это приводит к синтезу адаптивных ферментов, которые расщепляют лактозу до глюкозы и галактозы. Активация транскрипции структурных генов оперона называется индукцией; индуктором (активатором транскрипции) в данном случае является сама лактоза. Таким образом, ген-регулятор через белок-репрессор осуществляет индукцию генов оперона на уровне транскрипции (негативный контроль регуляции: транскрипция осуществляется в отсутствии регуляторного белка).
У эукариот во всех клетках многоклеточного организма содержится одинаковая ДНК, различие между клетками обусловлено тем, какие гены в них экспрессируются и какие белки в них синтезируются. Гены общеклеточных функций экспрессированы постоянно, а гены специализированных функций регулируются. Регуляция экспрессии генов происходит на уровне всех матричных процессов: претранскрипционный (спирализация и деспирализация хроматина, метилирование цитозинов в определенных сайтах молекулы ДНК и др.), транскрипционный (различная степень инициации процесса), посттранскрипционный (различные варианты процессинга, отбор в цитоплазме зрелых м-РНК для трансляции на рибосомах), трансляционный (изменение интенсивности синтеза белка на стадии инициации), посттрансляционный (после синтеза полипептида активация, инактивация или компартментализация молекул белка).
Для большинства генов важен контроль на уровне транскрипции.
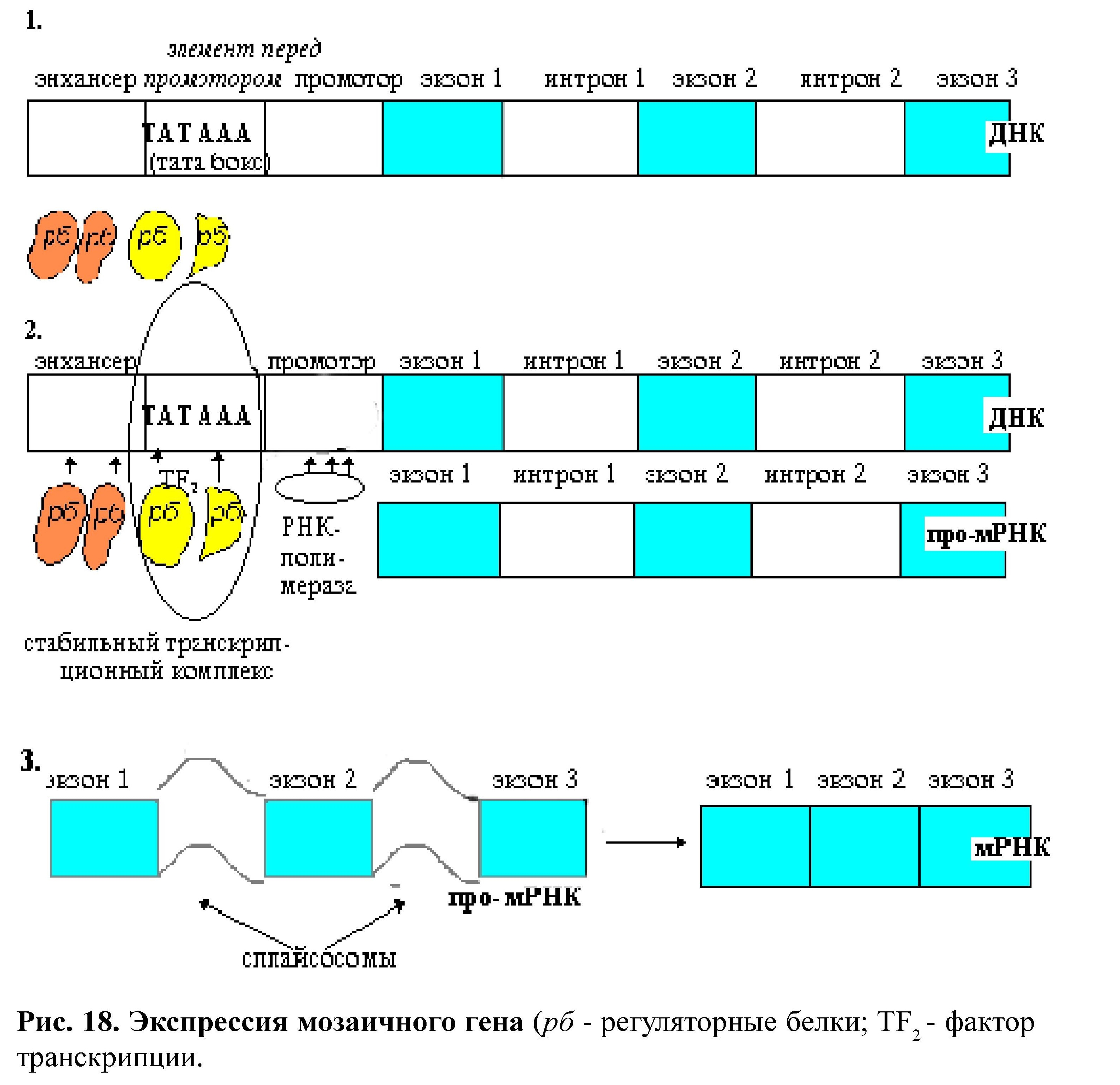
Строение генов (мозаичные), наличие регуляторных последовательностей (энхансер, сайленсер и элемент перед промотором), специфические последовательности промотора определяют особенности регуляции экспрессии генов на уровне транскрипции (рис. 18) .
- сложная инициация транскрипции:
для «узнавания» промотора и присоединения к нему РНК-полимеразы необходимо образование стабильного транскрипционного комплекса (соединение ТАТА-бокса с ТАТА-фактором ТF2).
- регулирование скорости и интенсивности транскрипции:
регуляторные белки (сайт - специфические ДНК связывающие) взаимодействуют с короткими нуклеотидными последовательностями в области энхансера и элемента перед промотором. Комбинационный эффект регуляторных белков (+ - ) обеспечивает экспрессию гена.
- наличие сплайсинга: (образование мРНК).
экзоны сплайсируются, сплайсинг катализируется сплайсосомами. Они узнают определенные последовательности интронов и удаляют их с образованием лассоподобных структур. В результате сплайсинга образуется мРНК, которая намного меньше, чем сам мозаичный ген.
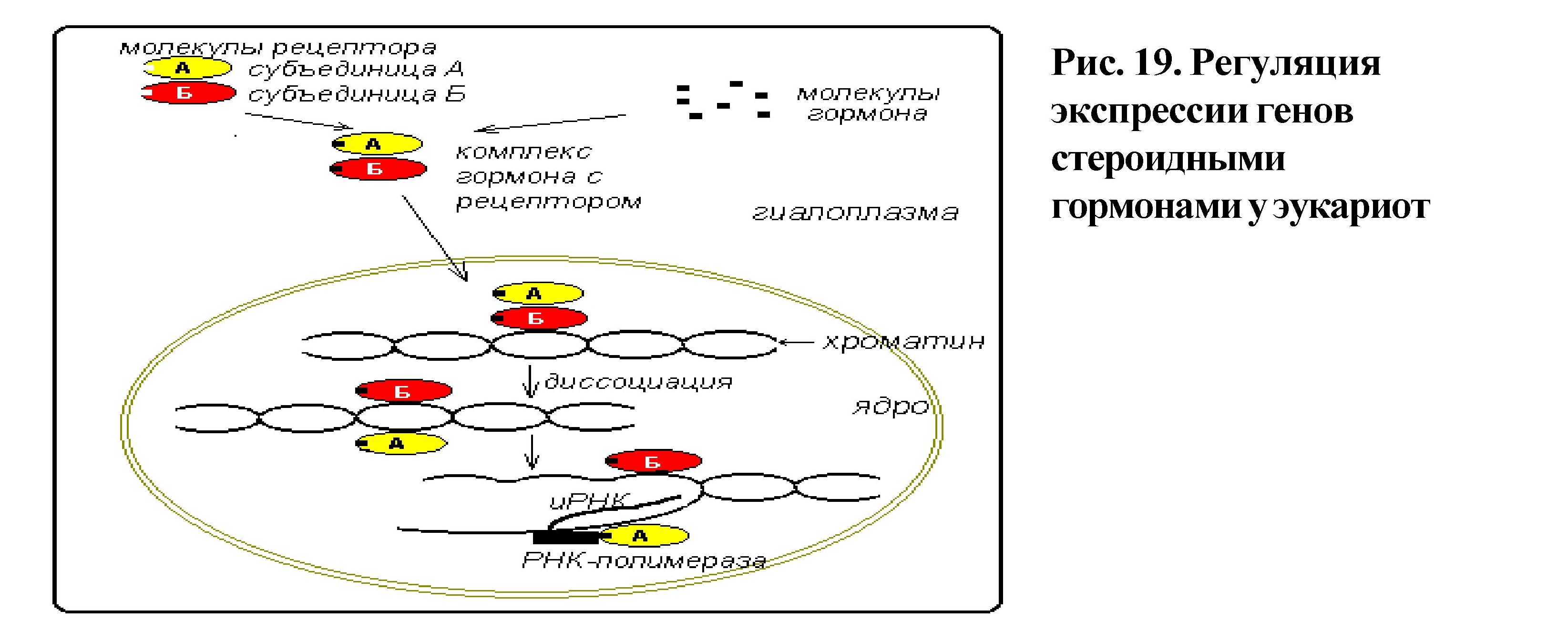
Изучена регуляция на уровне транскрипции при действии стероидного гормона прогестерона (рис. 19). В цитоплазме клеток-мишеней имеется белок-рецептор для гормона, который состоит из двух субъединиц А и В. Молекулы
гормона, проникая в цитоплазму клеток, связываются с рецептором, образуя комплекс рецептор-гормон. Комплекс перемещается из цитоплазмы в ядро, при этом В-субъединица в составе комплекса соединяется со специфическими участками хроматина и происходит диссоциация субъединиц. А-субъединица прикрепляется к ДНК в месте локализации соответствующего промотора, что изменяет активность РНК-полимеразы и приводит к увеличению синтеза мРНК и белка. Результатом действия гормона является усиление метаболических процессов, происходящих в клетке, а не инициация новых. Транскрипция генов происходит в присутствии активного регуляторного белка (позитивный контроль). Например, отсутствие белка-рецептора для гормона тестостерона приводит к формированию синдрома Морриса.
У эукариот, наряду с регуляторными процессами, влияющими на жизненный цикл клетки, имеются процессы, которые определяют развитие всего организма. К таковым относится генетический контроль дерепрессии генов глобина на разных стадиях эмбриогенеза - включение и выключение генов А и В кластеров, детерминирующих синтез глобиновых цепей и гемоглобина в целом. Гены глобина располагаются в А и В кластерах и экспрессируются в определённой последовательности слева направо (рис.20).
У человека образование «взрослого» гемоглобина происходит путём последовательной дерепрессии генов и включает три стадии: эмбриональный гемоглобин, гемоглобин плода и гемоглобин взрослого. Эмбриональный гемоглобин (до 8 недель развития) состоит из двух идентичных -подобных и двух идентичных -подобных цепей: две (дзета)- и две (эпсилон)-цепи, каждая из которых сходна с полипептидной цепью взрослого организма и позднее заменяется на неё. Гемоглобин плода включает две -цепи и две (гамма)-цепи. «Взрослый» гемоглобин состоит из двух цепей и двух цепей, а также (дельта)-цепи. У взрослого человека 97% всего гемоглобина составляет гемоглобин 2 2, 2% - гемоглобин 2 2, 1% - представлен гемоглобином плода 2 2.
Итак:
- гены, детерминирующие синтез глобина, дерепрессируются постепенно и в такой последовательности, как они располагаются в кластере;
- синтез «взрослого» гемоглобина обеспечивается постепенным включением и выключением генов А и В кластеров и синтезом соответствующих и глобиновых полипептидных цепей. Первой синтезируется -подобная цепь (дзета), которая заменяется на саму -цепь. Преобразование -подобных цепей происходит более сложно: первыми синтезируются (эпсилон)- и (гамма)-цепи, которые позднее заменяются на (дельта)- и -цепи.
- на разных этапах онтогенеза у человека имеются разные типы гемоглобина: эмбриональный, плодный и постнатальный.
- синтез глобиновых цепей происходит в различных органах: вначале в желточном мешке, затем в печени и селезёнке, а позднее и постоянно в костном мозге;
- все глобиновые гены сходны по своей функциональной активности.
Примером регуляции генов на уровне организма является и смена синтеза а-фетопротеина в постэмбриональном периоде на сывороточный альбумин.
III. Строение мозаичного гена
В конце 70-х годов было выяснено, что у эукариот имеются гены, которые содержат «лишнюю» ДНК, не представленную в молекуле мРНК. Они получили название мозаичных, прерывистых генов; генов, имеющих экзон-интронное строение.
1.Мозаичные гены эукариот имеют больший размер, чем последовательность нуклеотидов, представленная в мРНК (3-5%).
2.Мозаичные гены состоят из экзонов и интронов. Интроны удаляются из первичного транскрипта и отсутствуют в зрелой мРНК, которая состоит только из экзонов. Число и размеры интронов и экзонов индивидуальны для каждого гена, но интроны по размерам значительно больше экзонов.
3.Ген начинается экзоном и заканчивается экзоном, но внутри гена может быть любой набор интронов (гены глобина имеют 3 экзона и 2 интрона) (рис. 20). Экзоны и интроны обозначаются цифрами или буквами в порядке их расположения вдоль гена.).
4. Порядок расположения экзонов в гене совпадает с их расположением в мРНК.
5.На границе экзон-интрон имеется определённая постоянная последовательность нуклеотидов (ГТ - АГ), присутствующая во всех мозаичных генах.
6. Экзон одного гена может быть интроном другого.
7. В мозаичном гене иногда нет однозначного соответствия между геном и кодируемым им белком, то есть одна и та же последовательность ДНК может принимать участие в синтезе различных вариантов белка.
8. Один и тот же транскрипт (про-мРНК) может подвергаться разному сплайсированию, в результате этого сплайсированные участки мРНК могут кодировать разные варианты одного белка.
9. Особенности строения мозаичного гена позволяют осуществлять альтернативный сплайсинг (экзон L - экзон 2,3 или экзон S - экзон 2,3): синтезировать несколько вариантов белка на основе информации одного гена; создавать удачные комбинации белков, а если таковые неудачны, то производить отбор на уровне мРНК при сохранении неизменной ДНК (рис. 21).
В этом проявляется принцип экономного использования генетической информации, т.к. у млекопитающих в процессе транскрипции участвуют приблизительно 5-10% генов.
V. Эволюция представлений о гене
Проблема гена - универсальная проблема генетики. В процессе развития генетики менялись представления о гене: Г. Мендель, Т. Морган, Н.К. Кольцов, А.С.Серебровский, Н.П. Дубинин, С. Бензер, Дж. Бидл, Е. Тейтум, Эвери, Дж. Уотсон и Ф. Крик и др.
Современная теория гена базируется на крупных достижениях науки: установление сложной структуры гена, расшифровка генетического кода, генной инженерии и пр. В настоящее время ученые закончили работу над международным проектом «Геном человека», цель которого - полное определение первичной структуры геномной ДНК человека (3,5 млрд. нуклеотидных пар.). Реализация этого проекта позволила картировать, секвенированть и идентифицировать многие гены человека.
VI. Генная инженерия Генная инженерия - это совокупность методов получения генов и переноса генетической информации из одних организмов в другие. Целью генной иженерии является конструирование генетических структур, создание клеток (организмов) с новой генетической программой.
Методы генной инженерии включают следующие этапы:
- получение генетического материала (выделение природных генов, ферментативный или химический синтез генов).
- включение генов в векторную молекулу (плазмиды, вирусы, фаги) и создание рекомбинантной молекулы ДНК.
- введение рекомбинантных молекул ДНК в клетку-реципиент и их включение в хромосомный аппарат клетки.
- отбор трансформированных клеток, в геном которых включен переносимый ген.
- клонирование (размножение) клеток с рекомбинантной ДНК.