I. Определение индивидуального развития. Периодизация онтогенеза
Биология развития - область науки о живом, которая возникла на стыке эмбриологии, молекулярной биологии, генетики. Она изучает наследственные, молекулярные, структурные и функциональные основы индивидуального развития, механизмы регуляции жизнедеятельности организма. Онтогенез или индивидуальное развитие организма - это полный цикл развития особи, в основе которого лежит реализация наследственной информации на всех стадиях существования в определенных условиях среды. Это совокупность взаимосвязанных и детерминированных хронологических событий, закономерно совершающихся в процессе жизненного цикла. Филогенез - процесс исторического развития организмов. Онтогенез обусловлен филогенезом каждого вида. Онтогенез - это процесс непрерывный, однако, для решения конкретных научных и практических задач предложено несколько схем периодизации. В соответствии с одной из них выделяют 3 периода: преэмбриональный, эмбриональный, постэмбриональный. Преэмбриональный период (прогенез) соответствует гаметогенезу и оплодотворению, т.к. эти процессы во многом определяют развитие организма. С общебиологических позиций в соответствии со способностью организма осуществлять функцию полового размножения онтогенез делят на 3 периода: дорепродуктивный, репродуктивный, пострепродуктивный. В комплексе наук, составляющих биологию развития, ключевое место принадлежит эмбриологии, т.к. именно в зародышевый период онтогенеза происходит формирование структур и функций основных систем организма, реализуется программа наследственности.
II. Общие закономерности эмбрионального развития животных Эмбриология - наука, изучающая закономерности зародышевого развития организмов. Основы эмбриологии заложил петербургский академик К. Бэр (1792- 1876), который впервые описал яйцеклетку млекопитающих и показал, что зародыш позвоночных развивается вначале из двух зародышевых пластов. Основные работы в области сравнительной эмбриологии были выполнены русскими эмбриологами А.О. Ковалевским, И.И. Мечниковым, А.Н. Северцовым.
Эмбриональный или зародышевый период онтогенеза начинается с момента оплодотворения и продолжается до выхода зародыша из яйцевых оболочек. Относительно короткие отрезки эмбриогенеза, различаемые по характеру формообразовательных процессов, называют стадиями развития.
Собственно эмбриональное развитие включает стадии дробления, гаструляции, а также органо- и гистогенез. Процессы гаметогенеза и оплодотворения называют прогенезом. Цитологически они составляют промежуточное звено, связывающее онтогенезы родителей с онтогенезом их потомства. Преэмбриональный период (прогенез) включает развитие сперматозоидов и яйцеклеток из первичных половых клеток в процессе гаметогенеза. Морфофизиологические особенности яиц имеют важное значение для начальных фаз эмбриогенеза. Первичные половые клетки обособляются на ранних стадиях эмбриогенеза (из энтодермальных клеток - у анамний или из клеток желточного мешка - у амниот) и мигрируют в развивающиеся железы. У человека между 2-м и 5-м месяцами овогонии интенсивно размножаются, а к 7- му месяцу входят в профазу первого деления мейоза. Важным явлением в овогенезе является амплификация генов - это образование многочисленных копий генов, кодирующих синтез рРНК, необходимой для раннего эмбриогенеза. Многие хромосомы приобретают вид ламповых щеток, на них синтезируются впрок рРНК и мРНК - это период малого роста. В цитоплазме яйцеклетки происходит образование питательных веществ белка (желтка), гликогена, жира. Это сопровождается увеличением размеров яйцеклетки (период большого роста), что ведет к снижению ядерно-цитоплазменных отношений. Яйцеклетки, содержащие небольшое количество желтка, который распределен равномерно, называются изолецитальными. У большинства позвоночных желтка в цитоплазме много и он распределен неравномерно. Такие яйца называют телолецитальными. В цитоплазме яйцеклеток синтезируется набор специфических регуляторных белков: фактор дезинтеграции ядерной оболочки, фактор, вызывающий конден-сацию хромосом, фактор, преобразующий ядро сперматозоида в пронуклеус (активирующий в нем синтез ДНК перед дроблением), фактор, ответственный за цитотомию, фактор определяющий блок мейоза. В цитоплазме яйцеклеток под мембраной выделяется поверхностный кортикальный слой, толщиной 2-3 мкм, содержащий микрофиламенты и гранулы полисахаридной природы. На стадии накопления желтка в яйцах большинства животных выявляется полярность, отмечается анимально-вегетативная ориентация: количество желтка увеличивается по направлению к вегетативному полюсу. Отмечается ооплазматичская сегрегация - неравномерное распределение веществ цитоплазмы: мРНК, гликоген концентрируются на анимальном полюсе, аскорбиновая кислота - на экваторе, желток - на вегетативном полюсе. Благодаря этому уже на уровне яйцеклетки можно представить карту презумптивных зачатков - частей будущего организма. Этот процесс особенно активно продолжается после оплодотворения. Кроме плазматической мембраны яйца имеют дополнительные оболочки: первичную, вторичную, третичную. Оплодотворение - процесс слияния половых клеток, в результате которого образуется зигота. Он складывается из 3-х этапов:
1. Сближение гамет и проникновение сперматозоида в цитоплазму яйцеклетки. Гамета выделяет вещества - гамоны, которые активируют движение сперматозоида, происходит акросомная реакция - растворение яйцевых оболочек, слияние мембран яйцеклетки и сперматозоида. Образуется цитоплазматический мостик, ядро и центриоль сперматозоида входят в цитоплазму яйцеклетки.
2. Кортикальная реакция: изменяется кортикальный слой ооплазмы (кортикальные гранулы растворяются) и образуется оболочка оплодотворения (желточная). Перевителлиновое пространство обеспечивает блок полиспермии.
3. Активация метаболизма яйцеклетки характеризуется синтезом белка на уровне трансляции, т.к. мРНК, тРНК, рибосомы и энергия были запасены еще в овогенезе. Яйцеклетка в момент встречи со сперматозоидом находится на одной из стадии мейоза (метафазы II). Блок мейоза снимается, мейоз завершается, после чего ядро яйцеклетки превращается в женский пронуклеус. Ядро сперматозоида принимает вид профазного, в нем удваивается ДНК и формируется мужской пронуклеус. Оба пронуклеуса сближаются и сливаются, образуя общую метафазную пластинку - это сингамия. Первое митотическое деление приводит к образованию 2-х клеток (бластомеров) с набором хромосом 2n 2с.
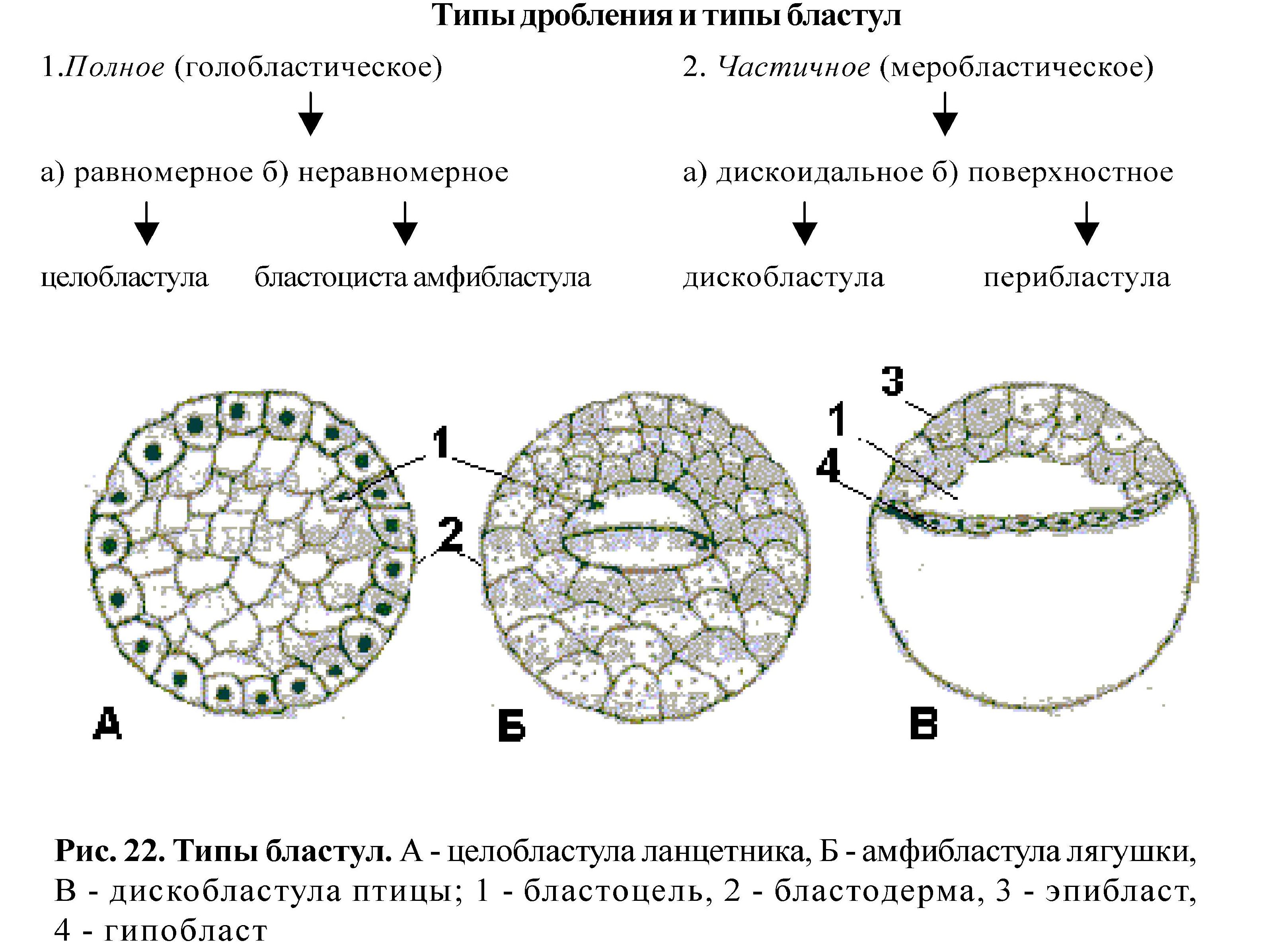
Эмбриональное развитие начинается процессом дробления, который состоит из последовательных митотических делений зиготы и далее бластомеров. Процесс дробления заканчивается образованием многоклеточного зародыша - бластулы. Сначала бластомеры плотно прилегают друг к другу, образуя морулу, затем между клетками образуется полость, бластомеры оттесняются к периферии, образуя стенку бластулы - бластодерму, а полость внутри - бластоцель. Порядок и способ дробления зависит от строения яиц, от количества и распределения желтка. Тип дробления определяется правилом Сакса-Гертвига: ядро стремится расположиться в центре свободной от желтка цитоплазмы, а веретено деления - в направлении наибольшей протяженности этой зоны.
По характеру взаиморасположения бластомеров и скорости деления различают: радиальное дробление (у иглокожих), билатеральнное (у аскариды), спиральное (у моллюсков), анархичное (у медузы).
Особенности молекулярно-генетических и биохимических процессов при дроблении.
1. Митотические циклы укорочены (у морского ежа - 30-40 мин).
2. В ДНК бластомеров больше точек инициации и синтез ДНК идет во всех репликонах одновременно.
3. Транскрипция генов и синтез иРНК начинается на разных стадиях развития. Если в яйцеклетке много желтка и других веществ - на стадии ранней бластулы. У млекопитающих - на стадии 2-х бластомеров: при этом синтезируются белки клеточных мембран, ферменты, необходимые для деления. Важная роль принадлежит цитотомии. Борозды дробления проходят по границам между участками ооплазмы, отражающими явление ооплазматической сегрегации, и цитоплазма разных бластомеров различается по химическому составу.
Гаструляция - превращение однослойного зародыша бластулы в многослойный (2-х- или 3-х слойный), называемый гаструлой. Бластодерма преобразуется в наружный зародышевый листок -эктодерму и внутренний - энтодерму, которая формирует полость внутри - гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. Наличие 2-х зародышевых листков у всех многоклеточных - свидетельство гомологии и единства происхождения животного мира. Развитие третьего зародышевого листка - мезодеомы -является эволюционным усложнением фазы гаструляции. Выделяют 4 разновидности направленных в пространстве перемещений клеток в процессе гаструляции:
1. Инвагинация - впячивание участка бластодермы внутрь целым пластом.
2. Эпиболия - обрастание мелкими клетками анимального полюса более крупных, отстающих в скорости деления клеток вегетативного полюса.
3. Деламинация - расслоение клеток бластодермы на 2 слоя, лежащих друг над другом.
4. Иммиграция - перемещение групп или отдельных клеток. В каждом случае эмбриогенеза сочетаются несколько способов гаструляции.
Мезодерма - третий зародышевый листок образуется двумя способами: телобластическим или энтероцельным.
На стадии гаструлы начинается цитодифференцировка, т.е. активное использование генетической информации собственного генома. Одним из регуляторов генетической активности является различный химический состав цитоплазмы клеток зародыша вследствие ооплазматической сегрегации.
Стадия гисто- органогенеза - образование тканей и органов. Зародышевые листки, занимая определенное положение по отношению друг к другу, обеспечивают взаимодействие между клеточными группами - эмбриональную индукцию. При этом обособляются клеточные группы, изменяется форма, структура и химический состав клеток. Появляются зачатки будущих органов. Процессы морфогенеза сопровождаются дифференциацией клеток, образованием тканей и избирательным и неравномерным ростом отдельных органов и частей организма.
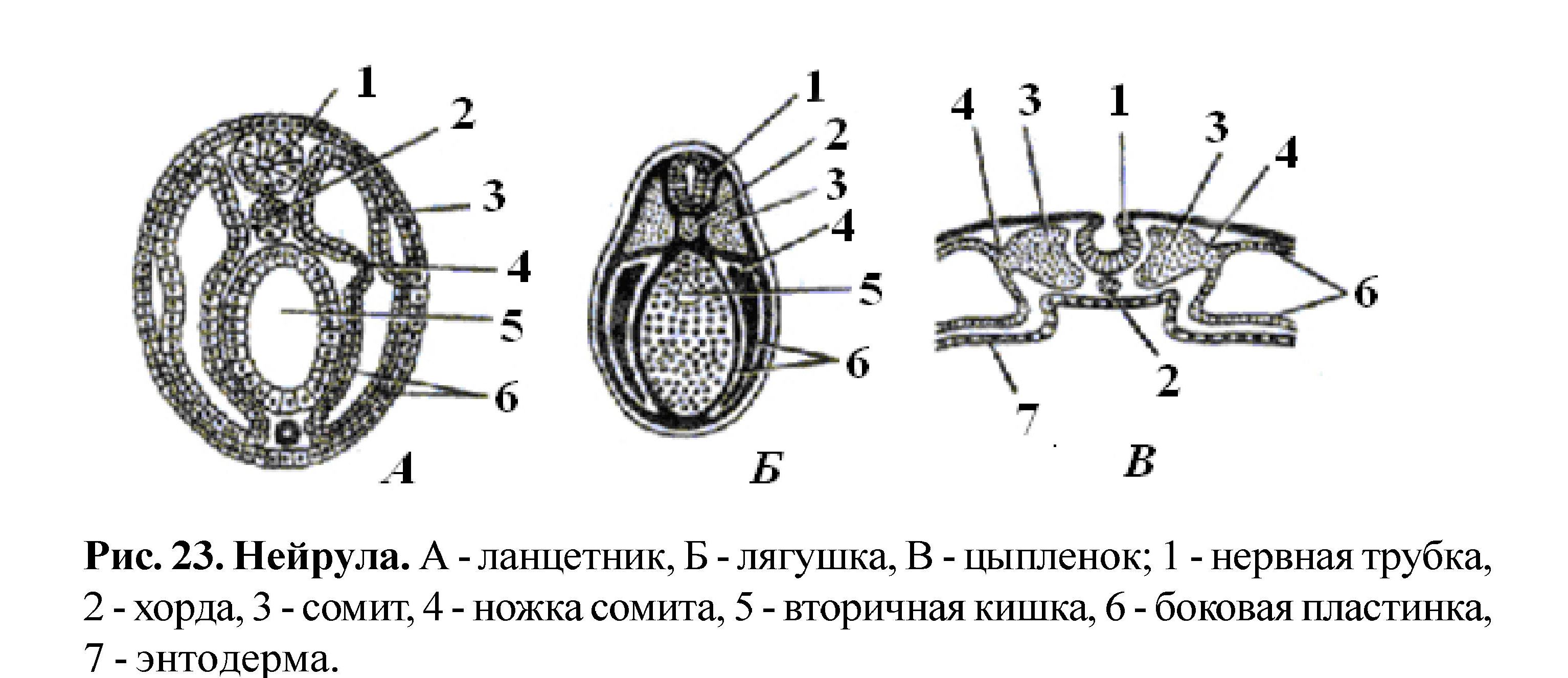
Начальную стадию органогенеза называют нейруляцией (рис. 23), она характеризуется формированием комплекса осевых органов: нервной пластинки, замыканием ее в нервную трубку, а также формированием хорды и вторичной кишки. Образование комплекса осевых органов - нервной трубки, хорды, кишки - характерная черта организации всех хордовых. Мезодерма по бокам от хорды расщепляется на парные сегменты - сомиты. Экто- мезо- и энтодерма в ходе дальнейшего развития, взаимодействуя друг с другом, участвуют в формировании определенных органов.
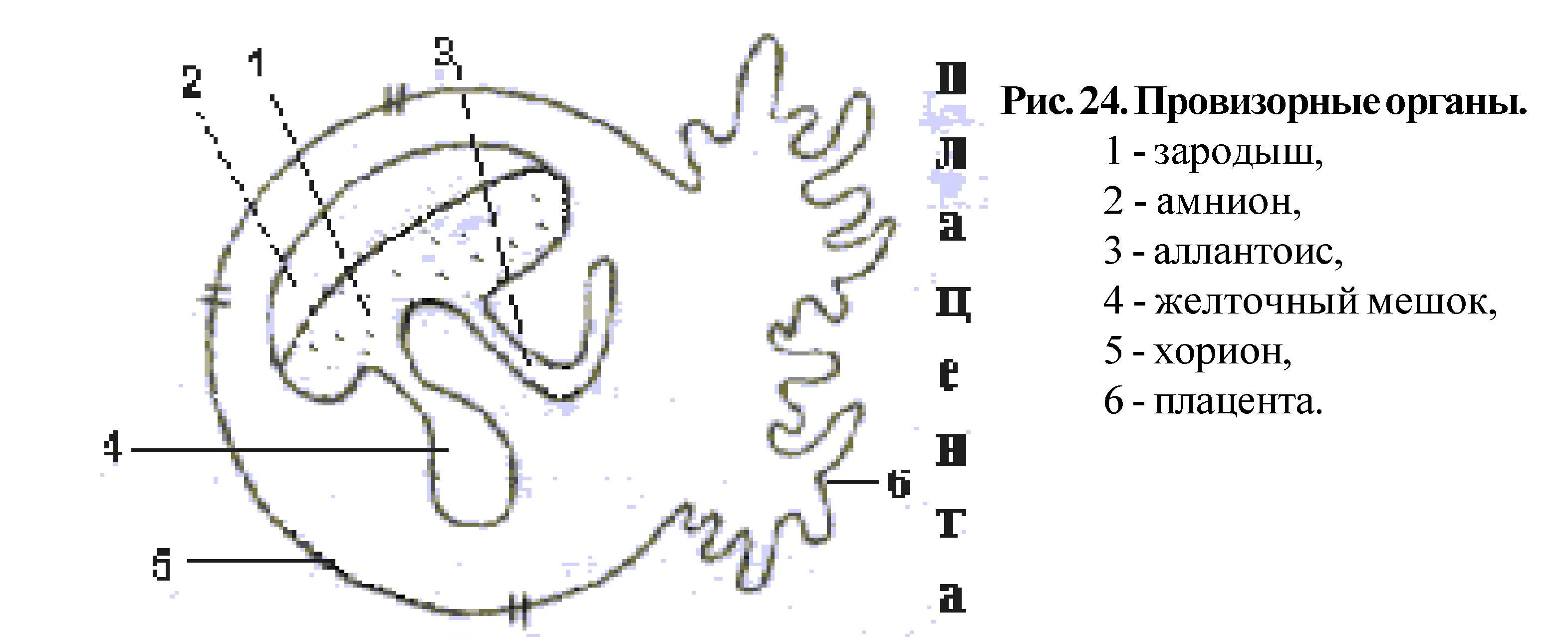
У большинства позвоночных к провизорным органам относятся амнион, хорион, желточный мешок и аллантоис, у плацентарных - еще и плацента.. Наличие или отсутствие амниона позволяет разделить позвоночных на 2 группы. Анамнии - эволюционно более древние - круглоротые, рыбы, земноводные - их развитие происходит в водной среде.
К группе Амниот относят первичноназемных; пресмыкающихся, птиц и млекопитающих. Амнион и хорион развиваются из эктодермы, желточный мешок - из энтодермы, аллантоис из энтодермы и спланхноплевры.
III. Механизмы онтогенеза
Выявление механизмов клеточных и системных процессов пространственных и временных преобразований в жизненном цикле организма составляет основную проблему современной биологии развития. Современная биология развития рассматривает следующие механизмы онтогенеза: 1. Деление клеток играет важную роль: 1) из одноклеточной стадии развивается многоклеточный организм; 2) деление обеспечивает рост структур организма; 3) обеспечиваются морфогенетические процессы в эмбриогенезе, а у взрослых - различные формы регенерации. Интерфаза имеет различную продолжительность в зависимости от стадии развития, локализации и функции клеток. Установлено, что многие структуры зародыша развиваются из небольшого числа клеток. Совокупность клеток, являющихся потомками одной родоначальной клетки называют клоном. Механизм отбора клеток неясен. На мышах показано, что на стадии бластоцисты в 64 клетки, дальнейшее развитие организма происходит из трех клеток. Известны мутации у дрозофилы, которые изменяют размеры организма за счет одного дополнительного деления. Т.о., клеточное деление протекает с разной интенсивностью, в разных частях, в разное время, носит локальный характер и подвержено мутационным изменениям. 2. Миграция клеток. Перемещения клеток имеют большое значение, начиная с процесса гаструляции. Клетки мезенхимного типа мигрируют одиночно или группами, а эпителия - пластом. Клетки нервного гребня, мигрируя, образуют два потока: поверхностный - включается в эпидермис, где дифференцируется в пигментные клетки; второй поток мигрирует в брюшном направлении, образуя чувствительные спинномозговые, симпатические и парасимпатические ганглии. Первичные половые клетки перемещаются из желточной энтодермы в зачаток половой железы. Нарушение миграции ведет к недоразвитию органов, нарушению нормальной локализации и возникновению пороков развития. Например, нарушение миграции нейробластов ведет к микрогирии и полигирии - аномальному расположению извилин в коре больших полушарий. Механизм миграции - амебоидные движения клеток по типу хемотаксиса или вследствие контактных взаимодействий со структурированным субстратом. 3. Сортировка клеток. Клетки не только перемещаются, но и узнают друг друга. Клетки экто-, энто- и мезодермы сегрегируют, собираются в группы. Это явление объясняется адгезией - избирательной слипаемостью клеток одного типа вследствие различий в поверхностном заряде мембран. Контактные взаимодействия основаны также на антигенных свойствах мембран. 4. Гибель клеток. Яркие примеры разрушений связаны с метаморфозом земноводных, насекомых. У человека закладка ребер у 7-го шейного позвонка подвергается некрозу. 5. Эмбриональная индукция - это взаимодействие частей развивающегося зародыша, когда один участок влияет на судьбу другого (опыты немецкого ученого Шпемана в 1924 г. на зародышах амфибий). Эффект индуцирующего воздействия определяется способностью воспринимать воздействие и отвечать на него, т. е. компетенцией. 6. Дифференцировка клеток - процесс приобретения специализации. Клетка приобретает химические, морфологические, функциональные особенности. Первые химические и морфогенетические различия между клетками обнаруживаются в период гаструляции. Зародышевые листки - пример ранней дифференцировки. Таким образом клетки, обладающие одинаковым кариотипом и генотипом дифференцируются в определенном направлении, соответственно виду организмов. В настоящее время общепринятой является точка зрения о дифференциальной экспрессии генов как основного механизма цитодифференцировки. Экспрессия гена в признак - сложный процесс, который можно изучать по продуктам активности генов биохимическим методом, электронно-микроскопически и др. Изучение политенных хромосом показало, что клетки разного типа содержат разные пуффы, т.е. транскрибируются разные гены. Видоизменяются и другие этапы экспрессии гена, происходят посттранскрипционные преобразования РНК, трансляция, посттрансляционные процессы. Важным в дифференцировке клеток является альтернативный процессинг первичных транскриптов.